(تابع. الجزء السابق: "الأتمتة من الحمض النووي والبروتينات: ما الذي تشترك فيه مع الإلكترونيات؟" )استقرنا على حقيقة أن معامل اللاكتوز مشابه للعنصر AND المنطقي. ولكن من أين تأتي خصائصه الرقمية؟ بعد كل شيء ، فإن إشارات الإدخال (تركيزات cAMP واللاكتوز) هي في الواقع تناظرية. دعنا نحاول رسم وظائف الإدخال لبروتين اللاكتوز.يتكون كل مدخل لمكبس اللاكتوز من مرحلتين من التعرف الجزيئي. يرتبط اللاكتوز بمكبس اللاكتوز ، ويربط المعوق بالحمض النووي. هناك عدة عشرات من جزيئات المعوق في الخلية ، كل منها ، طالما لا يوجد اللاكتوز ، يمكن أن يرتبط ببداية أوبرا اللاكتوز. عندما يكون مرتبطًا هناك ، لا يمكن أن يبدأ بوليميراز RNA في العمل. بسبب الحركة الحرارية لجزيء المعوق ، فإنه يستمر في السقوط من الحمض النووي ويرتبط مرة أخرى. إذا لم يكن هناك أي قمع على الإطلاق ، يعمل الأوبرون بكامل قوته (الآن للبساطة نعتبر أن هناك فائضًا من المنشط التقويمي). عند تركيزات منخفضة من المكثف ، فإنه يقلل من نشاط الأوبرون خطيا تقريبا. ولكن بعد ذلك يكون تأثير كل جزء جديد من المكثف أقل وأقل ، وبشكل عام ، يكون الرسم البياني للاعتماد قريبًا من المبالغة y = 1 / (x + 1)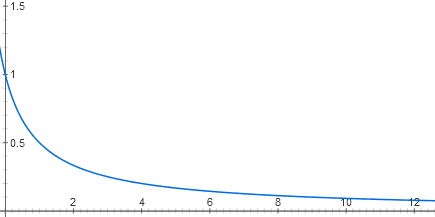
ماتان. . ?
, , . , :
A + B <-> C
, , - . , - ( , [A], [B] [C] ). : , , .
[C] / ([A] * [B]) = K
. (. . ) . , , .
, , , , ( ) . [C] ([A] * [B]) .
, : RepA + OpA <-> Op ( ). {R-O} = [Op] / ([OpA] * [RepA]). [] ( ) [RepA] ( ). , , : [Op] + [OpA] = 1
[Op] [OpA] :
{R-O} = (1 — [OpA]) / ([OpA] * [RepA])
:
[OpA] = 1 / (K{R-O} * [RepA] + 1)
y = 1 / (x + 1)
تطفو جزيئات المعوِّق في الخلية وترتبط بالحمض النووي في عبوات من أربعة فقط. لكي يصبح الشكل النشط للمثبط غير نشط ، يجب أن يرتبط بأربعة جزيئات من اللاكتوز - واحد أو اثنين ليس كافيًا ، فأنت بحاجة إلى جزيء واحد من اللاكتوز لكل وحدة فرعية بروتينية من المعاد رباعي. في حين أن هناك القليل من اللاكتوز في الخلية ، فإن جزيئاتها ترتبط بالمكبس واحد أو اثنين ولا تزال نشطة. ولكن بعد تركيز عتبة معينة من اللاكتوز ، فإن غالبية جزيئات الكابتون تربط أربعة جزيئات اللاكتوز وتنتقل إلى شكل غير نشط. وبالتالي فإن الرسم البياني لنشاط المكثف اعتمادًا على كمية اللاكتوز له شكل S ويتم وصفه بوظيفة الشكل y = 1 / (1 + x ^ 4).ماتان[RepA] [Lac]. , : .
RepA + 4 Lac <-> Rep
:
{R-L} = [Rep] / ([RepA] * [Lac]^4)
, :
[Rep] + [RepA] = n, n ≈ 50
, [RepA] = n / ({R-L} * [Lac]^4 + 1)
الآن ، من أجل الحصول على خاصية الإدخال من الأوبون لللاكتوز ، من الضروري استبدال الوظيفة الثانية في الأولى. سيكون لها الشكل ، حتى المعاملات ،y = 1 / (1 + (1 / (1 + x ^ 4)))جدولها الزمني هو أيضا على شكل حرف S. اتضح أن مُعامِل اللاكتوز لا يستجيب لتركيزات منخفضة من اللاكتوز. عندما يتم الوصول إلى تركيز العتبة ، تظهر جزيئات المثبط غير النشطة التي تربط أربعة لاكتوز لكل منها ، ويعمل الأوبرون بشكل حاد إلى حد ما.يتألف المدخل التنظيمي الثاني لبروتين اللاكتوز أيضًا من خطوتين للتعرّف الجزيئي: يرتبط cAMP بالمُنشط ، والمُنشط للحمض النووي. الفرق من مدخل اللاكتوز هو أن ربط الجزيئات (المنشط أو المخيم) هنا يزيد من نشاط ما اتصلوا به (الأوبرون أو المنشط). بمعنى ، ستنتقل الرسوم البيانية من الصفر وتقترب من الخط الأفقي في مكان ما فوق المحور X.ماتان, , :
2 + Akt <-> AktA
K{-} = [AktA] / ([Akt] * []^2)
, [AktA] [Akt] , 100 , . [Akt] [AktA], :
K{-} = [AktA] / ((100 — [AktA]) * []^2)
[AktA] = K{-} * []^2 / (1 + K{-} * []^2)
[AktA] [] y = x^2 / (1 + x^2)
نظرًا لربط جزيئين من cAMP ، لا يبدو الرسم البياني مثل hyperbole ، ولكن كمنحنى على شكل S. على الرغم من أن تأثير العتبة عليه أقل ملحوظة من الرسم البياني لللاكتوز ، لأن الدرجة ليست الرابعة ، ولكن الثانية فقط.اعتماد نشاط الأوبرون على كمية المنشط له فارق بسيط مختلف. إذا كان ربط المعوق يثبط الأوبرون بشكل شبه كامل (في مكان ما يصل إلى 0.1٪ ، وهو أقل من خطأ القياس البيوكيميائي) ، فإن غياب المنشط يمنع النشاط حتى 5٪ فقط. لذلك ، سيخرج المخطط ليس من الصفر ، ولكن من النقطة (0 ؛ 0.05):ص = 0.05 + (0.95 * س / (1 + س))ماتان« , » ,
= [OpA] + 0,05 * [Op]
[OpA] [AktA] . , , [AktA] . , , [AktA] , :
[OpA] = K{-O} * [AktA] / (K{-O} * [AktA] + 1)
y = x / (1 + x)
, ! , (1 — ) * 0,05. , , :
y = 0,05 + (0,95 * x / (1 + x))
يبقى للحصول على اعتماد نشاط الأوبرون على تركيز المخيم. للقيام بذلك ، استبدل الصيغة الأولى (لتركيز CAP النشط اعتمادًا على تركيز cAMP) في الصيغة الثانية (لنشاط الأوبرون من تركيز ATS النشط) ...... واحصل على جزء من أربعة طوابق:حصلنا على الوظيفة الأكثر تعقيدًا على الإطلاق في هذه المقالة. ومع ذلك ، فإن الرسم البياني الخاص به يشبه رسمًا بيانيًا يعتمد بشكل أبسط على CAP النشط على تركيز cAMP ، y = x ^ 2 / (1 + x ^ 2):وهو أيضًا منحنى على شكل حرف S مع بعض تأثير العتبة ، والذي يقترب ببطء من الأفقي y = 1. يبدأ فقط من الصفر ، ولكن من النقطة (0 ؛ 0.05).يبقى أن نفهم كيف يتفاعل المدخلان. في حالة مُعامِل اللاكتوز ، فإن الجواب بسيط - لا يمكن. لا يؤثر مكبّر اللاكتوز ومُنشّض الأيض على ارتباط بعضها ببعض بالحمض النووي. لذلك ، يمكن اعتبار ربط هذين البروتينين أحداثًا مستقلة. يتم تحقيق النشاط الكامل للأوبرون عندما يكون كل من المنشط متصلاً ولا يكون المكثف موصلاً. احتمال مثل هذه المصادفة يساوي ناتج احتمالات كل منها على حدة. لذا ، للحصول على دالة النشاط لأوبرون اللاكتوز من متغيرين ([Lac]) و ([cAMP]) ، ما عليك سوى مضاعفة وظائف كل من هذا المتغير:النشاط = (1 / (1 + (1 / (1 + [Lac] ^ 4))))) (0.05 + 0.095 * ([cAMP] ^ 2 / (1 + [cAMP] ^ 2)) / ( 1 + ([cAMP] ^ 2 / (1 + [cAMP] ^ 2)))))رسم بياني ثلاثي الأبعاد لهذه الوظيفةيبدو أن هضبة تنتهي في الوديان على جانبين. الخانق الأيمن (تركيزات منخفضة من اللاكتوز) أعمق وأكثر قاعدة مسطحة من اليسار (تركيزات cAMP منخفضة).في الحياة الواقعية ، لا يوجد E. coli من حوله إما لاكتوز على الإطلاق (في أغلب الأحيان) ، أو أن تركيزه أعلى من العتبة ولا يتداخل مكثف اللاكتوز عمليًا مع عملية الأوبرون (عندما أكل المالك شيئًا من منتجات الألبان). تركيز cAMP هو إشارة داخلية تنتجها الخلية نفسها. كما أنه إما صغير جدًا بحيث لا يتضمن بارون اللاكتوز (عندما يكون هناك جلوكوز أو نشا) ، أو يكفي لتشغيل 95٪ أو أكثر (إذا لم يكن هناك شيء ألذ من اللاكتوز). بمعنى ، دائمًا ما يكون أوبر اللاكتوز في حالة هضبة على هذا الرسم البياني ، أو في أحد الوديان.يتم قياس وظائف الإدخال لهرمون اللاكتوز تجريبيا. تشبه طريقة القياس تقنية تصحيح الميكروكنترولر "تعليق LED على الساق الإخراج." يتم أخذ المنطقة التنظيمية من الأوبرون وربطها بجين بروتين الفلوريسنت الأخضر. يتم إدخال هذا التكوين الجيني في خلايا Escherichia coli ، التي يتم من خلالها إزالة أوبرا اللاكتوز العادي (بحيث يتم ضبط تركيز اللاكتوز بدقة من قبل المجرب) والنظام القياسي الذي يرمز الجوع بمستويات cAMP. بعد ذلك ، يمكن قياس نشاط أوبرا اللاكتوز بدقة بواسطة مقياس الطيف الضوئي وفقًا لمستوى الفلورة الخضراء. وتبين أنه في حدود دقة القياس (1٪) ، تتقارب النظرية تمامًا مع التجربة ).الجزء السفلي من الخانق الأيسر على الرسم البياني لوظيفة الإدخال (بتركيزات منخفضة من cAMP) أعلى من الصفر. هذه ليست حشرة ، ولكنها ميزة: بفضل ذلك ، وبوجود العديد من السكريات في وقت واحد ، فإن E. coli مستعدة للتحول بسرعة إلى اللاكتوز عند انتهاء الجلوكوز. طفرات Escherichia coli ، التي تكون فيها الوظيفة التنظيمية لأوبرون اللاكتوز أقرب إلى نقية AND (أي أن الأوبرون لا يعمل بدون cAMP) ، يتحول من الجلوكوز إلى اللاكتوز لمدة ساعة أو أكثر: في وقت استنفاد الجلوكوز ، ليس لديهم إنزيمات امتصاص اللاكتوز ، مما يعني أنه لا يوجد الطاقة لجعلها بسرعة. تلتقي الإشريكية القولونية الطبيعية ("النوع البري") بهذه النقطة ، حيث تحتوي بالفعل على بعض الإنزيمات لامتصاص اللاكتوز ويستغرق التحول من السكر الجديد تمامًا 15-20 دقيقة.الآن دعنا نتطرق من أوبرا اللاكتوز ونلقي نظرة أكثر شمولاً على ما يمكن للعناصر المنطقية في مثل هذه القاعدة الكيميائية أن تفعل. في كل مكان تقريبًا في أنظمة الإشارات البيوكيميائية نجد التعرف الجزيئي ، والذي يتم وصفه من خلال وظائف النموذج y = x ^ n / (1 + x ^ n) للمنشطات و y = 1 / (1 + x ^ n) للمثبطات. توضح الدرجة n في هذه الوظائف عدد الجزيئات من نفس النوع التي يجب ربطها للحصول على تأثير ، وعادة ما تكون مساوية لـ 1 أو 2 أو 4 (في الغالب 2). يمكن الحصول على مفاتيح منطقية أكثر تعقيدًا عن طريق إضافة المزيد من مواقع ربط البروتين التنظيمي إلى بداية المعمل. يمكن أن يكون هناك أكثر من نوعين من هذه البروتينات. يمكن أن تضيف تأثيرات البروتينات المختلفة على هبوط البلمرة RNA (OR) أو تضاعف (AND).ولكن بشكل عام ، سنتعامل مع بعض التوليفة (الجمع أو الضرب) للوظائف الأساسية y = x ^ n / (1 + x ^ n) و y = 1 / (1 + x ^ n) بدرجات ومعاملات مختلفة.بعض الميزات المثيرة للاهتمام بسيطة للغاية. على سبيل المثال ، يمكن أن يرتبط نفس البروتين التنظيمي (في شكل مزدوج) بمنطقتين من المروج. في قسم واحد ، يعمل كمقمع ، وفي قسم آخر - كمنشط. لكي يعمل الأوبرون ، من الضروري أن يكون موقع المعوق فارغًا وأن موقع المنشط مشغول. اتضح وظيفة بحد أقصى واضح:y = (1 / (1 + x ^ 2)) * (x ^ 2 / (1 + x ^ 2))بدون هذا البروتين ، يكون المعامل غير نشط لأن موقع المنشط فارغ ، وبتركيزات عالية يكون غير نشط لأن الموقع مشغول المكبّر. سيكون النشاط الأقصى عندما يكون كل من موقع المنشط وموقع المعوق مشغولين باحتمال 50٪.لا يزال بإمكانك إنشاء موقعين ملزمين بالمنشط ، في أحدهما يرتبط المنشط بقوة بالحمض النووي ، ولكنه ينشط الجين بشكل ضعيف ، وفي الآخر - يرتبط ضعيفًا بالحمض النووي ، ولكنه ينشط الجين بقوة. في هذه الحالة ، نحصل على دالة مثل y = x ^ 4 / (1 + x ^ 4) + 0.3 * (10x) ^ 4 / (1 + (10x) ^ 4) ، مع رسم بياني متدرج:الرياضيات ، عاي! هل يمكنك إخبارنا في التعليقات بما هو ممكن وما الذي لا يمكن الحصول عليه عن طريق إضافة وضرب هذه الوظائف الأساسية y = x ^ n / (1 + x ^ n) و y = 1 / (1 + x ^ n)؟في الجزء التالي ، سنتعامل مع دوائر هذه العناصر المنطقية.