
Seit Tausenden von Jahren sucht die Menschheit in den Himmel und möchte wie Vögel werden. Viele große Köpfe, von Leonardo da Vinci bis zu den Gebrüdern Wright, entwarfen eine Vielzahl von Flugzeugen, von denen einige auf dem Papier blieben, während andere die Vorfahren moderner Flugzeuge und Hubschrauber wurden. Jetzt ist es nicht schwer für eine Person zu fliegen - ich habe ein Flugticket und Voila gekauft. Die meisten von uns denken nicht einmal darüber nach, wie dieser riesige Metallvogel funktioniert, der uns von einem Kontinent auf einen anderen bringen kann. Die Wahrheit zwischen modernen Flugzeugen und echten Vögeln ist nicht so sehr gemeinsam, aber mit der Entwicklung autonomer Roboter ist die Situation etwas anders. Viele Wissenschaftler versuchen, einen Robotervogel zu erschaffen. Damit diese Initiative erfolgreich ist, muss man nicht nur verstehen, wie Vögel fliegen, sondern auch wie man landet. Heute werden wir mit Ihnen eine Studie treffen, in der Absolventen der Stanford University den Mechanismus zum Pflanzen eines Papageis namens Gary und seiner Verwandten eingehend untersuchten. Wie landen Vögel, wie verstehen sie, dass sie auf einer bestimmten Oberfläche landen können oder nicht, und wie ist die Kinematik ihrer Gliedmaßen während der Landung? Antworten auf diese und andere Fragen finden wir im Bericht der Forscher. Lass uns gehen.
Studienbasis
Was ist der Unterschied zwischen modernen Flugzeugen und Vögeln in Bezug auf die Landung? Die Boeing 777 benötigt beispielsweise eine Landebahn mit einer Länge von mindestens 2500 m, ein Hubschrauber benötigt eine flache (vorzugsweise) Oberfläche wie Hubschrauber. Vögel wiederum sind in der Geometrie der Pflanzfläche nicht beschränkt und können auf fast jeder Oberfläche landen (wenn wir die Vögel als Ganzes und nicht als eine einzige Art betrachten). Wenn wir über autonome Roboter sprechen, sind ihre Landefähigkeiten sehr begrenzt. Einige von ihnen können erfolgreich auf speziell entworfenen Oberflächen landen, aber dies kann nicht mit einem gewöhnlichen Specht verglichen werden, der auf vertikalen, geneigten, horizontalen Oberflächen landen kann, solange Krallen zum Festhalten vorhanden sind.
 Es ist bequem für mich (Ameisenspecht / Melanerpes formicivorus).
Es ist bequem für mich (Ameisenspecht / Melanerpes formicivorus).Die Forscher stellen fest, dass ihre Vorgänger bereits einige Studien durchgeführt haben, in denen die Dynamik der Beine und Flügel von Vögeln während des Pflanzens quantifiziert wurde. Es bleibt jedoch unklar, wie sich diese Dynamik in Abhängigkeit von den verschiedenen Arten von Oberflächen ändert, die als Landeplatz dienen. Schließlich ist nicht nur die Geometrie des Landebereichs wichtig, sondern auch die Oberflächentextur.
Es ist auch interessant, dass frühere Studien nicht an lebenden Vögeln durchgeführt wurden, weshalb es in der Praxis nicht möglich war, die Griffstärke der Pfoten und Krallen vollständig zu bewerten. Aus diesem Grund blieben einige Fragen offen: Wie genau verstehen die Vögel, dass der Landeplatz geeignet ist? Was ist die Dynamik der Gliedmaßen beim Berühren und Greifen? warum für Vögel, die auf einer Vielzahl von Oberflächen landen, eine Kleinigkeit ist, etc.
In der Arbeit, die wir heute betrachten, beschlossen die Forscher, die Mechanismen und die Dynamik der Landung von Baumvögeln durch Beobachtung der Papageien von Lesson (
Forpus coelestis ) zu testen.
 Wellensittich der Lektion.
Wellensittich der Lektion.Die Probanden landeten freiwillig auf Stangen mit verschiedenen Geometrien und Texturen. Insgesamt gab es 9 Optionen: 3 „natürlich“, 3 mit geändertem Durchmesser und 3 mit geänderter Textur.
"Natürliche" Optionen werden so genannt, weil die Sitzstangen aus echten Zweigen (19 mm Durchmesser) von drei verschiedenen Bäumen bestanden:
- Kalifornische Eiche - die Oberfläche ist an verschiedenen Stellen eines Astes rau;
- Ceiba ist großartig - die Oberfläche ist relativ glatt, die Zweige sind weich;
- duftender Osmanthus - die Oberfläche ist in Längsrichtung rau, Äste sind hart.
Für Experimente mit einem modifizierten Durchmesser des Pols wurden gewöhnliche Birkenstifte mit unterschiedlichen Durchmessern als Material verwendet:
- 38 mm - zu großer Durchmesser für diesen Papageientyp;
- 19 mm - der Papagei kann einen Teilgriff ausführen (vordere und hintere Finger berühren sich nicht);
- 6 mm - der Papagei kann einen vollen Griff ausführen (die vorderen und hinteren Finger berühren sich).
Für Experimente mit unterschiedlichen Texturen wurde ein Birkenpfahl mit einem Durchmesser von 19 mm in drei verschiedene Materialien eingewickelt:
- Schaum - weiche und elastische Textur;
- Teflon - rutschige Textur;
- Schleifpapier ist eine raue und raue Textur.
Der Papagei testet alle 9 Optionen für den Landeplatz.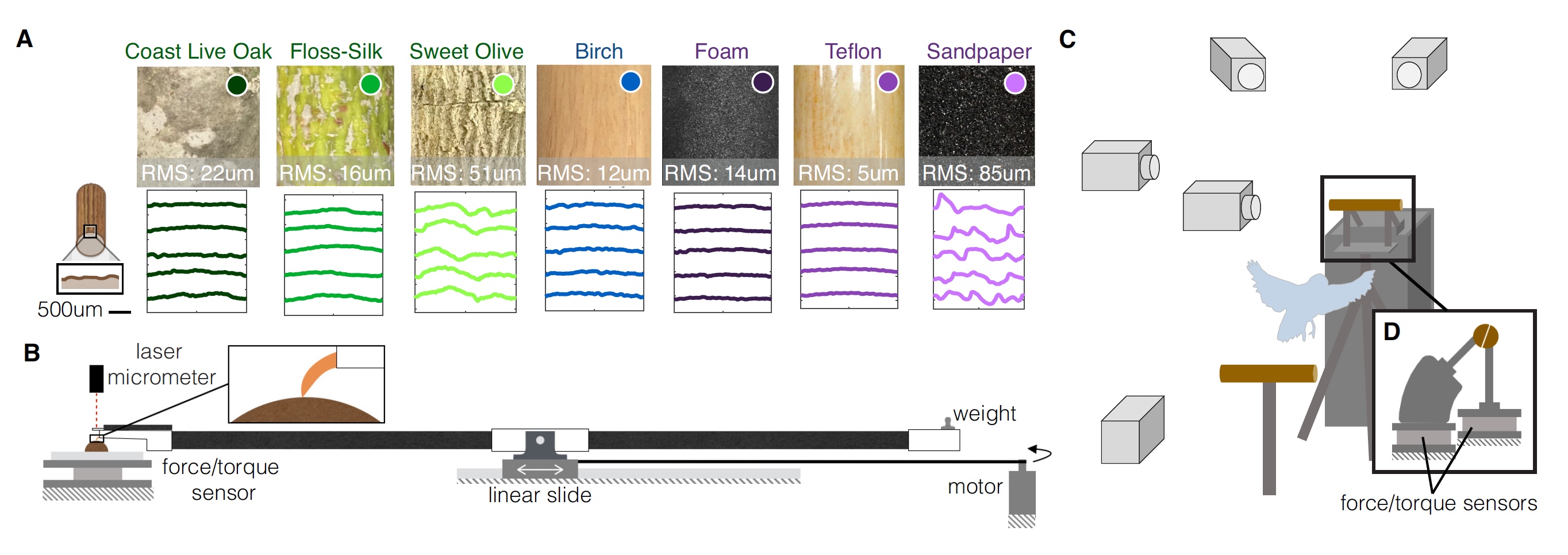 Bild Nr. 1
Bild Nr. 1Es wurde eine dreidimensionale Strukturlichtabtastung jeder der Perforationsvarianten durchgeführt, die es ermöglichte, die Oberflächentextur jeder von ihnen quantitativ zu beschreiben und Oberflächenprofile (
1A ) neu zu erstellen. Um die Auswirkung dieser Texturmerkmale auf das Zusammenspiel der Oberfläche von Barsch und Fuß des Papageien zu bewerten, führten die Forscher Widerstandstests an Fingern und Krallen durch, um die Reibungskräfte für jede Oberfläche zu messen. Der Klauendruck wurde auch bewertet, um die Oberflächenverformung (
1B ) zu messen.
In Diagramm
1C sehen wir, dass alle Tests unter dem Blick mehrerer Hochgeschwindigkeitskameras durchgeführt wurden und die Stangen in zwei Hälften zersägt wurden. Kraftsensoren und Kraftmomente sind an jeder Hälfte angebracht.
Forschungsergebnisse
Berücksichtigen Sie zunächst die Dynamik der Flügel und Beine während der Landung.
In allen Tests mit allen Varianten der Landefläche zeigten die Probanden die gleiche klassische Dynamik. Die Landung beginnt mit der Flügelbremsung (Luftphase), wonach die Pfoten den verbleibenden Impuls in Kontakt mit der Barsch absorbieren (Absorptionsphase). Der Kontakt wurde mit beiden Pfoten mit einer Zeitverzögerung von einigen Millisekunden zwischen ihnen hergestellt, d.h. Der erste Fuß in Kontakt mit der Stange ist der bevorzugte Fuß:
- Subjekt Nr. 1 - in 100% der Fälle die rechte Pfote;
- Proband Nr. 2 - in 83% der Fälle der linke Fuß;
- Proband Nr. 3 - in 85% der Fälle der linke Fuß.
Nach dem Kontakt beginnt die Befestigungsphase, wenn der Papagei seine Finger zuverlässiger und fester um die Barsch legt. Darauf folgt eine Phase, die nicht immer gefunden wird, aber häufig die Anpassungsphase ist, in der sie die Position der Pfoten oder Finger auf der Barsch irgendwie ändern.
Alle Phasen der Landung eines Papageis (entspricht dem Schema auf 2A ). Bild Nr. 2
Bild Nr. 2Manchmal flogen Papageien, die die Testbedingungen anscheinend überschätzten oder unterschätzten, leicht nicht über den Landeplatz (
2B ), was zu einer größeren Streuung des Neigungswinkels des Fußes führte, bei dem sie einen statischen Griff herstellten.
Es ist merkwürdig, dass die Durchschnittswerte der Landekraft für verschiedene Pole überraschend gleich bleiben, wenn die Indikatoren für die Stärke der Beine (
2B ) und die Richtung (
2C ) variieren.
Der Unterschied in den Indikatoren während der Landung kann durch die Papageienlandestrategie erklärt werden. Es gibt eine Theorie, nach der Vögel ihre Landung steuern, indem sie die Kontaktzeit mit der Oberfläche
τ (
t ) visuell beurteilen. Für eine erfolgreiche Landung passen sie ihre Annäherungsgeschwindigkeit an, um ein konstantes
τ (
t ) aufrechtzuerhalten. Wenn etwas detaillierter:
τ ist definiert als der Abstand zu den Barschen geteilt durch die Näherungsgeschwindigkeit (
v ).
Wenn der Vogel mit konstanter Verzögerung (
a ) langsamer wird, ist
τ =
s /
v = 0,5
bei 2 /
at = 0,5
t , in welchem Fall
τ (
t ) = 0,5.
Wenn
τ (
t ) <0,5 ist, nimmt die Hemmung bis zum Kontakt mit dem Barsch ab. Und wenn 0,5 <
τ (
t ) <1 ist, nimmt die Bremsung zu, bis der Vogel eine kontrollierte Kollision mit der Landefläche macht.
In Anbetracht der oben beschriebenen Kinematik halten Papageien relativ konstante Werte von
τ (
2E ) aufrecht, die Werten kontrollierter Kollisionen (
2F ) entsprechen. Der kleinste Mittelwert
τ = 0,80 wurde bei Tests mit einem Pol mit dem kleinsten Durchmesser (6 mm) beobachtet. Und der höchste Wert von
τ = 0,94 war bei Tests mit einer schaumbedeckten Stange. Dies deutet darauf hin, dass Papageien das Bremsen erhöhen, wenn sie sich einer weichen Landefläche nähern.
Das Vorhandensein eines hohen
τ (d. H. Größer als 0,5) in allen Tests mit allen Oberflächentypen legt nahe, dass die Vögel zum Zeitpunkt der Berührung die Landung nicht vollständig abgeschlossen haben.
Nachdem wir uns ein wenig mit der Dynamik der Landung vertraut gemacht haben, werden wir uns der Kinematik der Füße und Krallen unserer gefiederten Freunde zuwenden.
Angesichts der Gleichmäßigkeit des Verhaltens während der Landung, unabhängig von der Art der Oberfläche, kamen die Forscher zu dem Schluss, dass die Vögel auf Unterschiede in der Textur und Geometrie des Landepfostens genau mit ihren Pfoten und Krallen und nicht mit ihren Flügeln reagieren. Klingt logisch, nicht wahr? Überraschenderweise ist dies jedoch nicht ganz richtig.
Die Kinematik des Fußes zeigte einen ähnlichen stereotypen (für alle Arten von Sitzstangen gleichen) Satz von Landestufen (
3A ,
3B ).
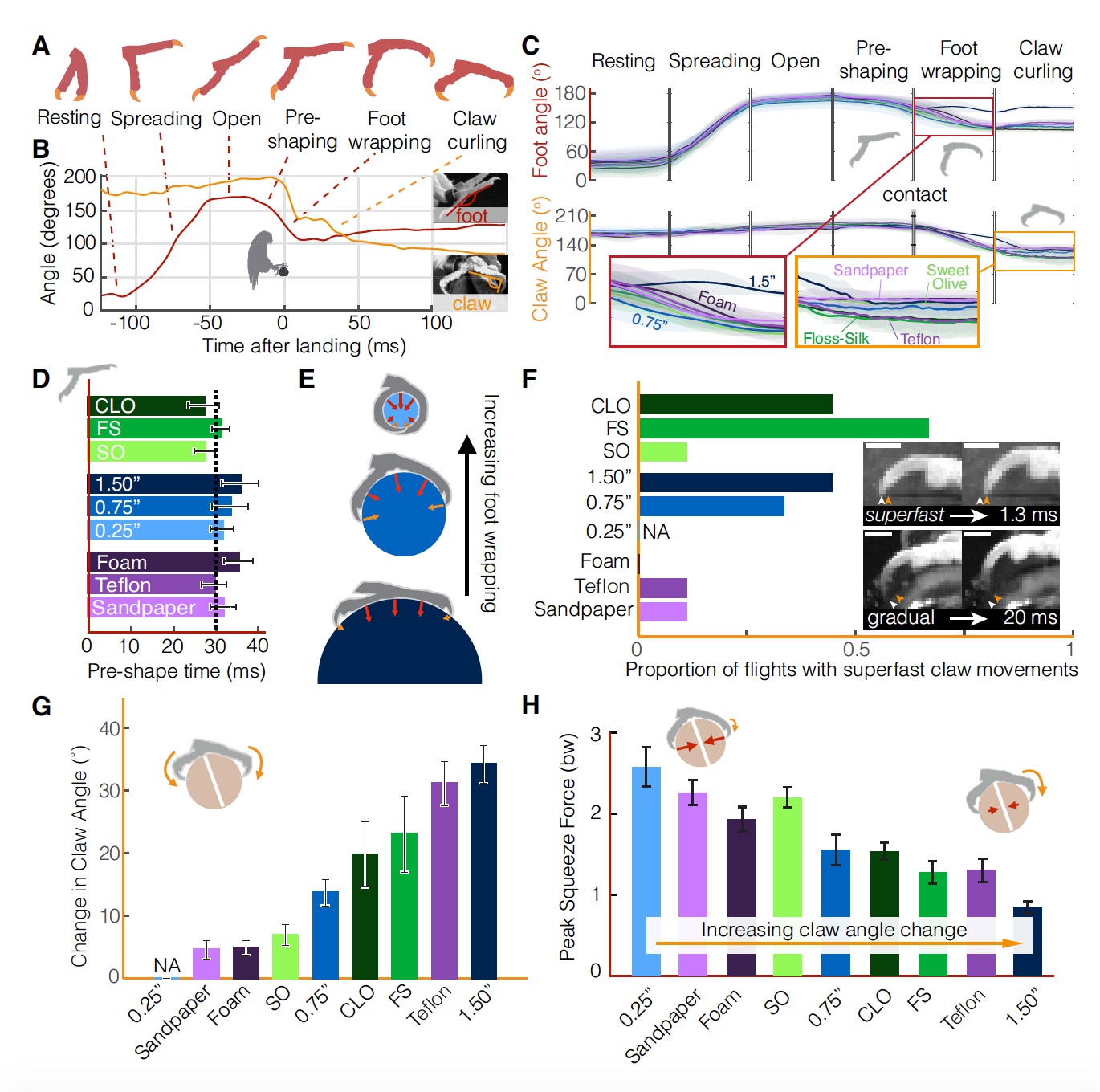 Bild Nr. 3
Bild Nr. 3Während des Fluges befinden sich die Füße in einem geschlossenen Zustand (Ruhephase), dann beginnen sie sich etwa 100 ms vor dem Berühren der Barsch zu öffnen (Öffnungsphase von 40 ± 8 ms), dann gibt es eine „offene“ Phase, wenn innerhalb von 21 ± 7 ms der Stopp erfolgt vollständig offenbart.
Unmittelbar vor dem Kontakt beginnen sich die Finger zu nähern (Phase der vorläufigen Bildung des Captures, Dauer 31 ± 10 ms). Nach dem Kontakt mit der Oberfläche beginnen die Finger, den Barsch zu erfassen (Umfangsphase, Dauer 19 ± 7 ms). Die letzte Phase ist die Klauenerfassung mit einer Dauer von 185 ± 11 ms.
Unterschiede in der Textur und Geometrie des Pols wirken sich ausschließlich auf die Kinematik der Landestufen nach Kontakt mit der Oberfläche aus (
3C ). Bis zum Kontakt bleibt die Kinematik von Fuß und Klaue in allen Tests unverändert, selbst wenn sich die Füße während der Vorgriffphase zu schließen beginnen.
Mit anderen Worten, wenn die Vögel auf den Unterschied in der Geometrie und Textur der Landefläche reagierten, würden kinematische Änderungen sogar während der Phasen vor dem Kontakt beobachtet, d. H. noch in der Luft. Stattdessen beginnen die Papageien 30 ms vor der Landung nacheinander mit der Vorerfassung (
3D ). Dies führt zu der Annahme, dass sie vorhersehbare Lösungen für das Problem der Landung auf einer ungewöhnlichen Oberfläche haben.
Es ist wichtig, dass der Vogel bis zum Moment des Kontakts einen großen Winkel zwischen den Krallen einhält, um deren Beschädigung zu verhindern. Ferner wird der Einfanggrad durch den Durchmesser des Pols (
3E ) bestimmt. Eine vollständige Erfassung erfolgt im Fall von Polen mit einem Durchmesser von 6 mm, d.h. Papageien können ihre Finger- und Krallenkraft voll ausnutzen, um sich zu stabilisieren.
Es ist wichtig zu beachten, dass im Moment des Kontakts die Krallen und / oder Pfoten aufgrund der glatten Oberfläche von den Stangen rutschen können. Infolgedessen kann der Klauenwinkel variieren. Wenn man bedenkt, dass die Klauengreifphase mehr als 100 ms dauern kann, können einzelne Bewegungen einer einzelnen Klaue (
3F ) unglaublich schnell (1-2 ms) erfolgen.
Superschnelle Klauenbewegungen werden wahrscheinlich nicht nur durch Muskelkontraktion reguliert, schlagen Forscher vor. Es ist auch überraschend, dass bei den Wirbeltieren die schnellsten Muskelbewegungen 5-10 mal langsamer sind als diese superschnellen Klauenbewegungen (zum Beispiel die menschlichen Armmuskeln - 50 ms, der Kolibri-Brustmuskel - 8 ms). Daraus können wir schließen, dass die Bewegungsgeschwindigkeit der Klaue eines Papageis von der Freisetzung der in der elastischen Sehne gespeicherten Energie und von der geringen Trägheit der Klaue abhängt. Solche schnellen Klauenbewegungen ermöglichen es Papageien, ihren Griff abhängig von der Geometrie und Textur der Landefläche sofort anzupassen.
Der Grad der Klauenerfassung hängt vom Durchmesser des Pols und von der Textur (
3G ) ab. Der ausgeprägteste Griff wird bei einem Durchmesser von 38 mm beobachtet. Es ist klarstellbar, dass es nur darum geht, die Krallen und nicht alle Beine vollständig zu erfassen. Das heißt, je größer der Durchmesser, desto mehr müssen Sie sich auf die Krallen verlassen, da die Pfoten nicht helfen (
3C ,
3E ). Der Klauenwinkel relativ zur Oberfläche hängt stark von seiner Textur ab. Beispielsweise „biegen“ sich die Krallen bei glatteren Oberflächen (Teflon) stärker (
3H ) als bei rauer Oberfläche (Sandpapier), da sie in diesem Fall die Oberfläche einfach etwas stärker komprimieren können und so eine zuverlässige Haftung gewährleisten. Übertrieben gesagt, biegen sich die Krallen stärker, wenn die Oberfläche schwerer zu komprimieren ist.
 Bild Nr. 4
Bild Nr. 4Anschließend beschlossen die Forscher zu überprüfen, wie sich aus mechanischer Sicht die Indikatoren für die Druckfestigkeit der Finger und die Biegung der Krallen ändern und wie sie sich auf die Anpassung des Vogels an die eine oder andere Oberfläche während der Landung auswirken.
Tests mit unterschiedlichen Texturen der Pole zeigten, dass sowohl die Fingerspitzen (
4A ) als auch die Krallen (
4C ) im Moment des Gleitens und Greifens eine ähnliche
Scherkraft * aufweisen .
Scherkraft * - ungleiche Kräfte, die einen Körperteil in eine bestimmte Richtung und den zweiten Körperteil in die entgegengesetzte Richtung drücken.
Für Pfotenpolster ist das Coulomb-Reibungsmodell (
4B ) die am besten geeignete Methode zur Darstellung der Scherkraft als Funktion der Normalkraft für alle Oberflächen, mit der die Richtung und Größe der Reibungskraft zwischen zwei Körpern in Kontakt mit trockenen Oberflächen vorhergesagt werden kann (daher wird dieses Modell auch als „Trockenreibungsmodell“ bezeichnet). )
Bei Krallen (
4C ) kann der Reibungskoeffizient 8-mal höher sein als der der Pfotenpolster (
4A ). Ein solcher Unterschied kann unter anderem durch die Ausrichtung der Klaue relativ zur Oberfläche (
4D ) verursacht werden: Auf der weichsten Oberfläche bleibt die Reibungskraft relativ konstant, auf einer rauen Oberfläche nimmt diese Kraft jedoch auch mit zunehmendem Klauenwinkel zu. Messungen zeigen jedoch auch bei gleichen Winkeln, dass der maximale Kraftkoeffizient für Krallen immer noch höher ist als für Beläge (
4E ).
Um die stochastische Natur der Kupplungshaftung auszugleichen, können Vögel den Lastausgleich und die Klauenbewegung auf der Oberfläche nutzen. Die Lastverteilung zwischen mehreren Krallen verringert effektiv die Verteilung der erwarteten Gesamtfestigkeit aller Krallen (
4E ). Darüber hinaus können Vögel ihre Krallen verlängern, wodurch sie die Wahrscheinlichkeit einer Adhäsion an einer unebenen Oberfläche erhöhen können, falls vorhanden. Dies erhöht den erwarteten Wert der Reibungskraft (
4F ), was die stärkere Biegung der Krallen auf Oberflächen erklärt, die schwieriger nur fest zusammenzudrücken sind (
3F ). Das Dehnen der Krallen bietet einen Vorteil bei Oberflächen, auf denen es nur wenige Unregelmäßigkeiten gibt, die jedoch recht groß sind. Diese Aussage steht jedoch leicht im Widerspruch zu den Beobachtungen, da die Papageien nicht sehr versucht haben, ihre Krallen zu verlängern, und dies legt nahe, dass sie während des Pflanzens nicht die besten Beulen auswählen. Daher biegen sie ihre Krallen so weit wie möglich, bis eine ausreichende Festigkeit erreicht ist, um einen stabilen Griff aufrechtzuerhalten. Wenn während dieses Vorgangs diese Kraft nicht erreicht wird, beginnt der Vogel, die Position der Krallen anzupassen oder abzuheben.
 Bild Nr. 5
Bild Nr. 5Der Unterschied zwischen den Reibungskoeffizienten für Krallen und Polster kann auch durch die Geometrie dieser Körperteile und die Geometrie der Landefläche erklärt werden, mit der sie in Kontakt stehen. Die Pads haben eine größere Fläche und eine weiche Textur, wodurch Sie die Last auf viele Kontaktpunkte verteilen können. Dies führt zu einer gleichmäßigeren Reibungskraft. Die Krallen sind wiederum grob und haben eine kleine Fläche, daher beruhen sie auf einer direkten Wechselwirkung mit der Geometrie der Oberfläche, d.h. auf Getriebe für Unebenheiten.
Um die geometrischen Effekte zu quantifizieren, charakterisierten die Wissenschaftler die Klauenform in der Sagittalebene in der Breite als Funktion der Höhe von der Klauenspitze (
5A ). Zusätzlich zur Verwendung vorhandener Oberflächenunregelmäßigkeiten können Krallen auch Reibungskräfte aus Oberflächenverformungen erzeugen.
Um diese Wechselwirkung zu simulieren, wurden Messungen der Eindringtiefe der Klaue in verschiedene Oberflächen (
5B ) und Messungen der Geometrie der Klaue selbst kombiniert. Die Geometrie der Klauenspitze wurde mit einer Kugel mit einem Radius von 50 μm für Lasten bis zu 25% des Körpergewichts (
5C ) modelliert. So wurde gefunden, dass zum Beispiel die Eindringtiefe in den Barsch aus kalifornischer Eiche 10 & mgr; m beträgt, d.h. 0,2% der Länge des äußeren Krallenbogens (ca. 5 mm).
Um die Auswirkung der Klauengröße auf die Traktion genauer zu bewerten, wurde ein anderes Modell mit Krallen mit unterschiedlichen Spitzendurchmessern erstellt, die an verschiedenen Oberflächen
haften (
5D ). Wie erwartet sind rauere Oberflächen ohne ein Eindringen von Klauen in die Oberfläche besser für eine erfolgreiche Erfassung geeignet (
5E ).
Wenn dem Modell Oberflächenpenetrationseffekte (
5F ) hinzugefügt wurden, wurde festgestellt, dass die nützliche Oberfläche erhöht war. Darüber hinaus ist der Winkel zwischen der vertikalen Achse und dem Nettokraftvektor auf die Klaue umso kleiner, je stärker der Effekt ist. Wenn der Radius und die Masse der Klauenspitze isometrisch skaliert werden und die ausgeübten Kräfte von der Masse abhängen, wird der Tiefenfaktor (Eindringtiefe geteilt durch den Radius der Klauenspitze) ebenfalls isometrisch skaliert.
Daraus können wir eine logische Schlussfolgerung ziehen, dass kleine Unregelmäßigkeiten auf dieser Oberfläche ausreichen, damit kleine Tiere effektiv an der Oberfläche haften.
 Bild Nr. 6
Bild Nr. 6Durch die Kombination aller oben genannten Modelle für verschiedene Effekte, Kräfte und andere Dinge erstellten die Forscher ein einziges zweidimensionales Modell, das gleichzeitig die Parameter von Krallen, Pfoten und dem Körper als Ganzes berücksichtigt (
6A ).
Die Gesamtheit der Daten zur Kontaktmechanik der Füße und der Landefläche ermöglichte eine begrenzte Optimierung, um den dreidimensionalen „Fangraum“ des Vogels und die notwendige Kombination von Kraft und Kraftmoment (
6B ) zu bestimmen. , , : , . , (
6C ).
, . , . , . : , .
, , . , , , . , .
.
Nachwort
, , . . , . , , , . , . - -, - — . , , - , . , , , .
Freitag off-top:
( , ), .
Vielen Dank für Ihre Aufmerksamkeit, bleiben Sie neugierig und wünschen Sie allen ein schönes Wochenende! :) :)
, . ? ? Unterstützen Sie uns, indem Sie eine Bestellung
aufgeben oder Ihren Freunden empfehlen, einen
Rabatt von 30% für Habr-Benutzer auf ein einzigartiges Analogon von Einstiegsservern, das wir für Sie erfunden haben: Die ganze Wahrheit über VPS (KVM) E5-2650 v4 (6 Kerne) 10 GB DDR4 240 GB SSD 1 Gbit / s von $ 20 oder wie teilt man den Server? ( RAID1 RAID10, 24 40GB DDR4).
Dell R730xd 2 ? 2 Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 $199 ! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — $99! . c Dell R730xd 5-2650 v4 9000 ?