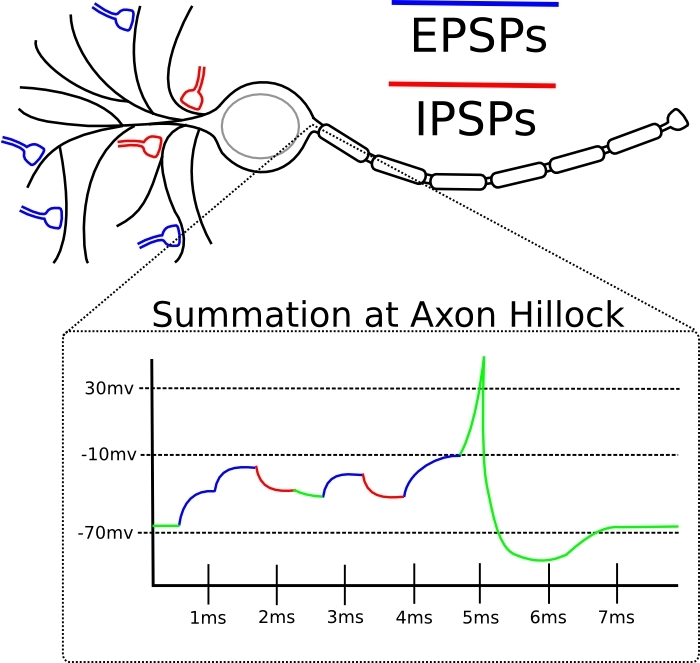
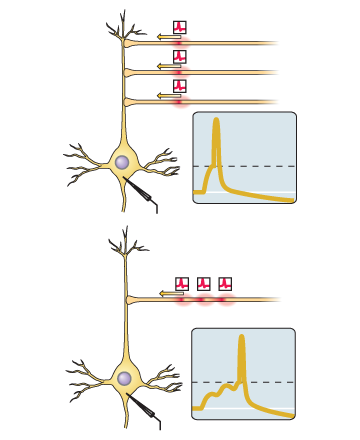
Si el pico de entrada a esta sinapsis tiende a aparecer justo antes de que la neurona misma genere el pico, entonces la sinapsis se amplifica.Si el pico de entrada a esta sinapsis tiende a ocurrir inmediatamente después de la generación del pico por la neurona misma, entonces la sinapsis se debilita.
Source: https://habr.com/ru/post/es390385/More articles:15 años del primer contacto del aparato de la Tierra con un asteroideEl número de usuarios de sitios y aplicaciones para citas en línea está creciendo rápidamenteUbuntu Tablet: "todo lo que necesitas de una PC está en una tableta"¿Trabajas demasiado? Las vacaciones no ayudarán14 de febrero - Día de Lawrence: Regalos para tu ser queridoLos titulares de derechos inventan nuevas formas de convertir a los piratas en usuarios que paganTecnología FRAMCalidad de comunicación: aplicación de Android del Ministerio de ComunicacionesCómo despertar juegos de interés en la historia o la mesa redonda de WargamingEl troyano bancario inteligente le permite retirar una cantidad casi ilimitada de dinero en cajeros automáticosAll Articles