Automatización a partir de ADN y proteínas: ¿qué tiene en común con la electrónica?
(respuesta del biólogo al artículo "No escribes animales correctamente" )Hay miles de genes en cada célula. Nunca es que todos trabajen simultáneamente. Las células musculares, hepáticas y cerebrales transportan el mismo conjunto de genes. Son muy diferentes porque diferentes genes funcionan en ellos. Todas las células pueden activar y desactivar genes individuales en respuesta a diversas influencias externas. Es decir, el sistema que controla la actividad de los genes es un sistema de control automático. Los científicos quieren entender cómo funciona esa automatización para poder repararla y descifrarla. Por ejemplo, en las células humanas hay un sistema de aproximadamente trescientos genes y proteínas que controla la división celular. Cuando se descompone y la célula comienza a dividirse todo el tiempo, se produce cáncer.Un diagrama simplificado de la red de genes que regula la división de las células humanas.¿Cómo es la automatización intracelular?
En el "hierro" se suele utilizar impulsos eléctricos. Las señales en biología suelen ser químicas, es decir, cambios en la concentración de cualquier sustancia. Hay señales eléctricas en el sistema nervioso, pero este es solo un dispositivo para la transmisión rápida a larga distancia, similar a la fibra óptica en tecnología. La transmisión de un impulso nervioso de una célula a otra se produce en forma química, memoria a largo plazo y la integración compleja de señales en una neurona también es química.Donde haya señales, habrá pistas: las señales están fuera de lugar y el ruido es basura aleatoria que se adhiere a las señales. En electrónica, la llegada de señales a la dirección es proporcionada por cables y aislamiento. La diafonía ocurre debido al acoplamiento capacitivo e inductivo parásito entre conductores. En una célula bacteriana, su contenido se mezcla más o menos uniformemente, y cualquier señal química (y hay miles de ellas) está disponible en cualquiera de sus puntos. Las células de animales y plantas se dividen en compartimentos con diferente composición química, pero los tipos de estos compartimentos son menos de una docena y cada uno contiene cientos y miles de sustancias diferentes.La llegada de la señal al lugar correcto de la célula se debe al reconocimiento molecular según el principio de "bloqueo de teclas". Una molécula de proteína puede reconocer otra proteína, una proteína puede reconocer una secuencia de ADN específica, una proteína puede reconocer moléculas pequeñas como los azúcares. Este reconocimiento no siempre es perfectamente preciso, por lo que las moléculas que tienen una forma similar a un compañero de reconocimiento regular crearán puntas. Además de la interferencia, hay ruidos en la electrónica. Surgen debido al movimiento térmico de los átomos, que introduce una fracción del caos en el movimiento de los electrones. En una celda, el ruido químico también está asociado con el movimiento térmico. Cuando la señal es débil, por ejemplo, diez piezas de moléculas de este tipo en toda la célula, estas moléculas se dispersarán alrededor de la célula al azar, y no el hecho de manera uniforme. Como resultado, su concentración en algún rincón de la célula fluctuará aleatoriamente, y esto será ruido.¿Qué se regula exactamente en una célula viva?
Una jaula puede considerarse como una planta que produce todo el equipo que necesita y puede ensamblar un segundo de la misma planta a partir de ella. El equipo principal son los ribosomas, máquinas CNC que recolectan todas las proteínas. La estructura de la proteína recolectada se registra en una cinta - matriz de ARN (ARNm). El ribosoma se mueve a lo largo del ARNm y, de acuerdo con sus instrucciones, recoge una cadena de una nueva proteína a partir de unidades individuales: aminoácidos. La cadena de proteínas que emerge del ribosoma se pliega en una bola compacta de proteína madura, que comienza a funcionar. Las proteínas pueden acelerar las reacciones químicas (enzimas), así como participar en el transporte de sustancias, señalización, defensa y muchas otras cosas.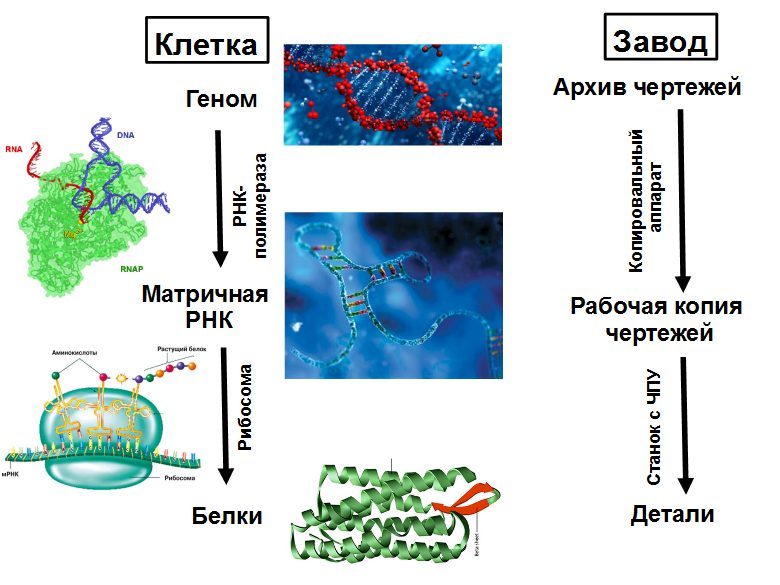
Elementos lógicos sobre ADN y proteínas.
Quizás el ejemplo más estudiado de un interruptor genético es el operón lactosa de Escherichia coli. El francés Jacob y Mono por su investigación recibieron el Premio Nobel en 1965. E. coli, como su nombre lo indica, vive en el intestino. Puede ser el intestino de una gran variedad de animales, desde abejas hasta humanos. Allí ella tiene que comer lo que le dio el dueño, para que pueda comer muchos nutrientes diferentes.E. coli bajo un microscopio y en una placa de Petri.E. coli puede crecer, por ejemplo, en un medio nutritivo que consiste solo en glucosa y sales minerales y producir todos los aminoácidos y vitaminas que necesita. En lugar de glucosa, puede haber otros azúcares (fructosa, azúcar de malta - maltosa, azúcar de leche - lactosa y una docena de otros), así como polisacáridos, como el almidón.Para absorber cada uno de estos azúcares, necesita sus propias enzimas. La producción de estas enzimas debe iniciarse solo cuando el azúcar correspondiente se encuentra en el medio ambiente; de lo contrario, se desperdiciarán los materiales y la energía para la producción de estas enzimas. Es decir, los genes de estas enzimas de digestión de azúcar deben activarse solo cuando este azúcar está presente. Para asimilar cada azúcar, por regla general, se necesitan varias enzimas, y no una. Sus genes están ubicados cerca de la cadena de ADN y su actividad está controlada por una región reguladora común de ADN al comienzo del primer gen. Tal grupo de genes colaborativos y controlados sincrónicamente se llama un "operón".Esquema del operón lactosa Eloperón lactosa consta de tres genes. El primero (LacZ) codifica una proteína de transporte que bombea lactosa a la célula, y los otros dos (LacI y LacA) codifican enzimas, cuyo trabajo conjunto convierte la lactosa en "azúcar predeterminado" - glucosa. Al comienzo del operón de lactosa hay un sitio de unión a ARN polimerasa (promotor) y un sitio de unión a proteínas reguladoras (operador).La actividad del operón lactosa está controlada por dos señales. La primera señal, obviamente, es la concentración de lactosa. Si no hay lactosa, entonces no se necesitan las enzimas de su asimilación. La segunda señal es un poco más complicada. En un experimento, Jacob y Mono cultivaron E. coli en un medio que contenía tanto lactosa como glucosa. Bajo tales condiciones, la barra primero consume glucosa, y su operón lactosa se apaga. Cuando termina la glucosa, el crecimiento de bacterias se detiene durante 15-20 minutos y luego continúa debido a la lactosa. Durante una pausa, el operón lactosa se enciende. Es decir, la segunda entrada reguladora apaga el operón de lactosa cuando la célula tiene más azúcares disponibles que la lactosa (la absorción de glucosa no requiere enzimas adicionales).Otros experimentos mostraron que el operón lactosa no responde a la glucosa en sí. En cambio, el nivel de hambre (más precisamente, la "bondad" de los azúcares disponibles) dentro de la célula de E. coli está codificado por una sustancia de señalización especial. Se llama "monofosfato de adenosina cíclico" o cAMP. Mientras el palo crece en glucosa, prácticamente no hay AMPc en la célula. Si no hay glucosa, pero hay maltosa o almidón (sustancias ligeramente más difíciles de asimilar), se produce un poco de AMPc. Si no hay maltosa, pero hay fructosa o lactosa, el nivel de AMPc en la célula será mayor. Si no hay azúcares, pero hay ácido láctico, es aún mayor, y finalmente, si no hay nada sabroso y tiene que comer glicerina, entonces el nivel de AMPc será el más alto. Entonces, dos señales controlan el operón de lactosa: concentración de lactosa y concentración de AMPc.¿Cómo afectan estas señales a la actividad genética? Un gen activo es aquel en el que la ARN polimerasa produce muchos ARN mensajeros. La ARN polimerasa siempre comienza a trabajar en este operón al aterrizar en la misma región de ADN (se llama el "promotor"). ¿Dónde está el lugar para lactosa y cAMP? La ARN polimerasa no puede reconocer todos los cientos de señales que regulan la actividad de miles de genes. Para esto, hay proteínas reguladoras especiales (se denominan "factores de transcripción"). Dos proteínas reguladoras están involucradas en el manejo del operón de lactosa, una para cada señal de entrada: un represor de lactosa y un activador catabólico.Represor de lactosa (púrpura) en el ADN Unrepresor de lactosa es una proteína que puede unirse a la lactosa o a una secuencia de ADN específica. Hay dos secuencias de este tipo en el genoma de Escherichia coli, ambas al comienzo del operón lactosa, a cada lado del promotor. Las moléculas represoras de lactosa se adhieren firmemente entre sí en grupos de cuatro moléculas de proteínas. La unión de tales cuatro al ADN retuerce la cadena de ADN en un circuito cerrado en el que termina el promotor. La ARN polimerasa no puede sentarse en un promotor tan retorcido y el gen se apaga. Si la lactosa aparece en la célula, se une al represor de lactosa y se cae del ADN, liberando el promotor de la ARN polimerasa: el operón comienza a funcionar. Así es como se organiza la entrada reguladora del operón según la concentración de lactosa.Un activador catabólico en el ADN Unactivador catabólico responde de manera similar a la concentración de AMPc. Es cierto que hay varias diferencias en los detalles:- el activador catabólico se encuentra en el ADN justo al lado del promotor y no se superpone. Por el contrario, mejora la unión de la ARN polimerasa al promotor y aumenta la actividad del operón.- un activador catabólico une cAMP y ADN. Sin AMPc, no puede unirse al ADN.- se necesita un activador catabólico para la actividad completa del operón lactosa, pero incluso sin él, el operón seguirá funcionando durante aproximadamente el 5% de la actividad total. El represor de lactosa regula el operón en el rango de 0 a 100%.- Las moléculas del activador catabólico están conectadas en pares, no en cuatro. En la próxima publicación veremos a qué conduce esta diferencia.Es decir, podemos escribir una tabla de verdad para el operón lactosa:El operón lactosa funciona casi como un elemento lógico AND. Solo el 5% de la actividad en el estado de "lactosa +, cAMP -" cae ligeramente de la imagen ideal.Continúa con gráficos y matan: https://geektimes.ru/post/283686/ Source: https://habr.com/ru/post/es399233/
All Articles