(continuación. Parte anterior: "Automatización a partir de ADN y proteínas: ¿qué tiene en común con la electrónica?" )Nos decidimos por el hecho de que el operón lactosa es similar al elemento lógico AND. ¿Pero de dónde vienen sus propiedades digitales? Después de todo, ambas señales de entrada (cAMP y concentraciones de lactosa) son, de hecho, analógicas. Intentemos dibujar las funciones de entrada del operón lactosa.Cada entrada del represor de lactosa consta de dos etapas de reconocimiento molecular. La lactosa se une al represor de lactosa, y el represor se une al ADN. Hay varias decenas de moléculas represoras en la célula, cada una de las cuales, siempre que no haya lactosa, puede unirse al comienzo del operón de lactosa. Cuando está unido allí, la ARN polimerasa no puede comenzar a funcionar. Debido al movimiento térmico de la molécula represora, se sigue cayendo del ADN y se vuelve a unir. Si no hay ningún represor, el operón funciona a plena potencia (ahora, por simplicidad, consideramos que hay un exceso de activador catabólico). A bajas concentraciones del represor, reduce la actividad del operón casi linealmente. Pero entonces el efecto de cada nueva porción del represor es cada vez menor, y en general, el gráfico de dependencia está cerca de la hipérbole y = 1 / (x + 1)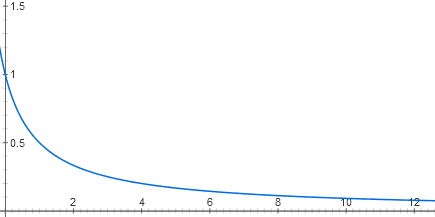
matan. . ?
, , . , :
A + B <-> C
, , - . , - ( , [A], [B] [C] ). : , , .
[C] / ([A] * [B]) = K
. (. . ) . , , .
, , , , ( ) . [C] ([A] * [B]) .
, : RepA + OpA <-> Op ( ). {R-O} = [Op] / ([OpA] * [RepA]). [] ( ) [RepA] ( ). , , : [Op] + [OpA] = 1
[Op] [OpA] :
{R-O} = (1 — [OpA]) / ([OpA] * [RepA])
:
[OpA] = 1 / (K{R-O} * [RepA] + 1)
y = 1 / (x + 1)
Las moléculas represoras flotan en la célula y se unen al ADN solo en paquetes de cuatro. Para que la forma activa del represor se vuelva inactiva, debe unirse a cuatro moléculas de lactosa: una o dos no son suficientes, necesita una molécula de lactosa para cada subunidad de proteína del represor cuádruple. Si bien hay poca lactosa en la célula, sus moléculas se unen al represor uno o dos y permanece activo. Pero después de un cierto umbral de concentración de lactosa, la mayoría de las moléculas represoras se unen a cuatro moléculas de lactosa y pasan a una forma inactiva. El gráfico de la actividad represora que depende de la cantidad de lactosa, por lo tanto, tiene forma de S y se describe mediante una función de la forma y = 1 / (1 + x ^ 4).Matan[RepA] [Lac]. , : .
RepA + 4 Lac <-> Rep
:
{R-L} = [Rep] / ([RepA] * [Lac]^4)
, :
[Rep] + [RepA] = n, n ≈ 50
, [RepA] = n / ({R-L} * [Lac]^4 + 1)
Ahora, para obtener la característica de entrada del operón para lactosa, es necesario sustituir la segunda función en la primera. Tendrá la forma, hasta coeficientes,y = 1 / (1 + (1 / (1 + x ^ 4)))Su horario también tiene forma de S. Resulta que el operón lactosa no responde a bajas concentraciones de lactosa. Cuando se alcanza el umbral de concentración, aparecen moléculas represoras inactivas que unen cuatro lactosas cada una, y el operón se activa de forma bastante brusca.La segunda entrada reguladora del operón lactosa también consta de dos pasos de reconocimiento molecular: el AMPc se une al activador y el activador al ADN. La diferencia con la entrada de lactosa es que aquí la unión de moléculas (activador o AMPc) aumenta la actividad de lo que contactaron (operón o activador). Es decir, los gráficos irán desde cero y se acercarán a la línea horizontal en algún lugar por encima del eje X.Matan, , :
2 + Akt <-> AktA
K{-} = [AktA] / ([Akt] * []^2)
, [AktA] [Akt] , 100 , . [Akt] [AktA], :
K{-} = [AktA] / ((100 — [AktA]) * []^2)
[AktA] = K{-} * []^2 / (1 + K{-} * []^2)
[AktA] [] y = x^2 / (1 + x^2)
Debido a la unión de dos moléculas de AMPc, el gráfico no parece una hipérbole, sino una curva en forma de S. Aunque el efecto umbral es menos notable que en el gráfico para la lactosa, porque el grado no es el cuarto, sino solo el segundo.La dependencia de la actividad del operón de la cantidad de activador tiene un matiz diferente. Si la unión del represor suprime el operón casi por completo (en algún lugar hasta el 0,1%, que es inferior al error de medición bioquímica), la ausencia de activador suprime la actividad solo hasta el 5%. Por lo tanto, el gráfico saldrá no desde cero, sino desde el punto (0; 0.05):y = 0.05 + (0.95 * x / (1 + x))Matan« , » ,
= [OpA] + 0,05 * [Op]
[OpA] [AktA] . , , [AktA] . , , [AktA] , :
[OpA] = K{-O} * [AktA] / (K{-O} * [AktA] + 1)
y = x / (1 + x)
, ! , (1 — ) * 0,05. , , :
y = 0,05 + (0,95 * x / (1 + x))
Queda por obtener la dependencia de la actividad del operón de la concentración de AMPc. Para hacer esto, sustituya la primera fórmula (por la concentración de CAP activa según la concentración de cAMP) en la segunda fórmula (por la actividad del operón a partir de la concentración de ATS activo) ...... y obtenga una fracción de cuatro pisos:Tenemos la función más compleja de todas en este artículo. Sin embargo, su gráfico se parece a un gráfico de una dependencia más simple del CAP activo de la concentración de cAMP, y = x ^ 2 / (1 + x ^ 2):También es una curva en forma de S con cierto efecto umbral, que se acerca lentamente a la horizontal y = 1. Solo comienza no desde cero, sino desde el punto (0; 0.05).Queda por entender cómo interactúan las dos entradas. En el caso del operón lactosa, la respuesta es simple: nada en absoluto. El represor de la lactosa y el activador catabólico no afectan la unión entre sí al ADN. Por lo tanto, la unión de estas dos proteínas puede considerarse eventos independientes. La actividad completa del operón se logra cuando tanto el activador está conectado como el represor no está conectado. La probabilidad de tal coincidencia es igual al producto de las probabilidades de cada uno de ellos por separado. Entonces, para obtener la función de actividad del operón lactosa de dos variables ([Lac]) y ([cAMP]), solo necesita multiplicar las funciones de cada una de estas variables:Actividad = (1 / (1 + (1 / (1 + [Lac] ^ 4))) * (0.05 + 0.095 * ([cAMP] ^ 2 / (1 + [cAMP] ^ 2)) / ( 1 + ([cAMP] ^ 2 / (1 + [cAMP] ^ 2))))Gráfico tridimensional de esta funciónParece una meseta que termina en gargantas en ambos lados. La garganta derecha (bajas concentraciones de lactosa) es más profunda y de fondo plano que la izquierda (bajas concentraciones de cAMP).En la vida real, E. coli a su alrededor no tiene lactosa (la mayoría de las veces), o su concentración es más alta que el umbral y el represor de lactosa prácticamente no interfiere con el funcionamiento del operón (cuando el dueño comió algo lácteo). La concentración de AMPc es una señal interna producida por la propia célula. También es demasiado pequeño para incluir el operón de lactosa (cuando hay glucosa o almidón), o lo suficiente como para activar el 95% o más (si no hay nada más sabroso que la lactosa). Es decir, casi siempre el operón lactosa se encuentra en las condiciones de una meseta en este gráfico o en una de las gargantas.Las funciones de entrada del operón lactosa se miden experimentalmente. El método de medición se asemeja a la técnica de depuración de microcontroladores "cuelgue el LED en la pata de salida". La región reguladora del operón se toma y se une al gen de la proteína fluorescente verde. Esta construcción génica se inserta en las células de Escherichia coli, de donde se extrae el operón de lactosa regular (para que el experimentador establezca estrictamente la concentración de lactosa) y el sistema estándar que codifica el hambre con niveles de AMPc. Después de eso, la actividad del operón lactosa se puede medir con precisión mediante un espectrofotómetro de acuerdo con el nivel de fluorescencia verde. Resulta que, dentro de los límites de precisión de medición (1%), la teoría converge completamente con el experimento ).La parte inferior del desfiladero izquierdo en el gráfico de la función de entrada (a bajas concentraciones de cAMP) está por encima de cero. Esto no es un error, sino una característica: gracias a él, en presencia de varios azúcares a la vez, E. coli está lista para cambiar rápidamente a lactosa cuando termina la glucosa. Los mutantes de Escherichia coli, en los que la función reguladora del operón de lactosa está más cerca de la Y pura (es decir, el operón no funciona sin AMPc), cambian de glucosa a lactosa durante una hora o más: en el momento del agotamiento de la glucosa, no tienen enzimas de absorción de lactosa, lo que significa que no hay energía para hacerlos rápidamente. La Escherichia coli normal ("tipo salvaje") cumple con este punto, ya que tiene algunas enzimas de asimilación de lactosa y se tarda 15-20 minutos en cambiar completamente a azúcar nuevo.Ahora divaguemos del operón de lactosa y veamos más ampliamente de lo que son capaces los elementos lógicos en una base química de este tipo. Casi en todas partes en los sistemas de señalización bioquímica encontramos el reconocimiento molecular, que se describe mediante funciones de la forma y = x ^ n / (1 + x ^ n) para los activadores y y = 1 / (1 + x ^ n) para los represores. El grado n en estas funciones muestra el número de moléculas del mismo tipo a unir para obtener un efecto, y generalmente es igual a 1, 2 o 4 (con mayor frecuencia 2). Se pueden obtener claves lógicas más complejas agregando más sitios reguladores de unión a proteínas al comienzo del operón. Puede haber más de dos tipos de estas proteínas. Los efectos de diferentes proteínas en el aterrizaje de la ARN polimerasa pueden sumar (OR) o multiplicarse (AND).Pero en general, trataremos con alguna combinación (suma o multiplicación) de las funciones básicas y = x ^ n / (1 + x ^ n) e y = 1 / (1 + x ^ n) con diferentes grados y coeficientes.Algunas características interesantes son muy simples. Por ejemplo, la misma proteína reguladora puede unirse (en forma emparejada) a dos regiones del promotor. En una sección, actúa como un represor, y en otra, como un activador. Para que el operón funcione, es necesario que el sitio represor esté vacío y que el sitio activador esté ocupado. Resulta una función con un máximo claro:y = (1 / (1 + x ^ 2)) * (x ^ 2 / (1 + x ^ 2))Sin esta proteína, el operón está inactivo porque el sitio activador está vacío, y en altas concentraciones está inactivo porque el sitio está ocupado represor La actividad máxima será cuando tanto el sitio del activador como el sitio del represor estén ocupados con una probabilidad del 50%.Todavía puede hacer dos sitios de unión para el activador, en uno el activador se unirá fuertemente al ADN, pero activará débilmente el gen, en el otro, se unirá débilmente al ADN, pero activará fuertemente el gen. En este caso, obtenemos una función como y = x ^ 4 / (1 + x ^ 4) + 0.3 * (10x) ^ 4 / (1 + (10x) ^ 4), con un gráfico escalonado:Matemáticas, ay! ¿Puede decirnos en los comentarios qué más es posible y qué no se puede obtener agregando y multiplicando estas funciones básicas y = x ^ n / (1 + x ^ n) e y = 1 / (1 + x ^ n)?En la siguiente parte, trataremos con circuitos de tales elementos lógicos.