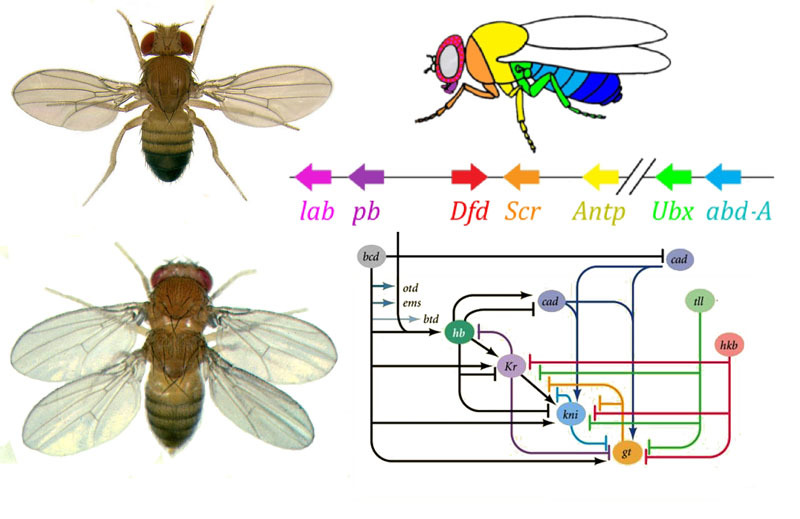
 ¿Cómo usar un huevo para convertir un huevo en una larva y luego en una mosca adulta? Una mosca, como otros insectos, consta de segmentos. Los segmentos se combinan en tres grupos: cabeza, pecho y abdomen. Un segmento puede tener un par de extremidades. Hay tres segmentos en el cofre, cada uno con un par de patas. El segundo segmento torácico también tiene alas, y el tercero, el escarabajo de tierra (pequeñas alas de cola, casi invisibles sin lupa). El abdomen consta de ocho segmentos; no hay extremidades en ellos. Inicialmente, había seis segmentos en la cabeza, pero durante la evolución de los insectos, los límites entre ellos se borraron por completo. Sus extremidades son antenas con las que la mosca huele y siente, y tres pares de extremidades bucales. Para un insecto o una cucaracha, serían las mandíbulas, los maxilares y el labio inferior, mientras que para una mosca todos se fusionan en la trompa.Para que todos los órganos estén en su lugar, cada célula debe saber dónde se encuentra en el embrión. Por lo tanto, en las primeras etapas del desarrollo embrionario, cuando todavía no hay órganos y todas las células se ven iguales, aparece una "rejilla de coordenadas" bioquímica con los ejes anteroposterior, dorsoabdominal e izquierdo-derecho. Las mutaciones de los genes que forman la cuadrícula de coordenadas pueden cambiar fácil y rápidamente la apariencia del animal: mover los órganos a otro lugar, cambiar su número, etc. Por lo tanto, tales genes son importantes para la evolución de los animales y se estudian muy activamente. La red de genes para marcar el embrión de una mosca Drosophila, el juguete favorito de los genetistas, es quizás la más estudiada entre las redes de genes animales.El algoritmo para marcar un huevo a lo largo del eje anteroposterior es básicamente simple:1) determinar qué extremo del huevo será la cabeza, qué cola2) dividir el embrión a lo largo del eje anteroposterior en 17 segmentos3) dividir cada segmento en la parte delantera, media y trasera4) marque la cabeza (6 segmentos), el cofre (3 segmentos) y el abdomen (8 segmentos)5) marque las diferencias individuales de los segmentos Lared de genes que se ocupa de esto tiene un dispositivo modular. Cada módulo realiza su propio paso del algoritmo general. La mosca Drosophila resultó ser un objeto muy conveniente para estudiar las redes de desarrollo de genes, ya que tiene deformidades hereditarias muy brillantes y reveladoras. Por ejemplo, Antennapedia mutación en la que las antenas se transforman en las piernas o los
¿Cómo usar un huevo para convertir un huevo en una larva y luego en una mosca adulta? Una mosca, como otros insectos, consta de segmentos. Los segmentos se combinan en tres grupos: cabeza, pecho y abdomen. Un segmento puede tener un par de extremidades. Hay tres segmentos en el cofre, cada uno con un par de patas. El segundo segmento torácico también tiene alas, y el tercero, el escarabajo de tierra (pequeñas alas de cola, casi invisibles sin lupa). El abdomen consta de ocho segmentos; no hay extremidades en ellos. Inicialmente, había seis segmentos en la cabeza, pero durante la evolución de los insectos, los límites entre ellos se borraron por completo. Sus extremidades son antenas con las que la mosca huele y siente, y tres pares de extremidades bucales. Para un insecto o una cucaracha, serían las mandíbulas, los maxilares y el labio inferior, mientras que para una mosca todos se fusionan en la trompa.Para que todos los órganos estén en su lugar, cada célula debe saber dónde se encuentra en el embrión. Por lo tanto, en las primeras etapas del desarrollo embrionario, cuando todavía no hay órganos y todas las células se ven iguales, aparece una "rejilla de coordenadas" bioquímica con los ejes anteroposterior, dorsoabdominal e izquierdo-derecho. Las mutaciones de los genes que forman la cuadrícula de coordenadas pueden cambiar fácil y rápidamente la apariencia del animal: mover los órganos a otro lugar, cambiar su número, etc. Por lo tanto, tales genes son importantes para la evolución de los animales y se estudian muy activamente. La red de genes para marcar el embrión de una mosca Drosophila, el juguete favorito de los genetistas, es quizás la más estudiada entre las redes de genes animales.El algoritmo para marcar un huevo a lo largo del eje anteroposterior es básicamente simple:1) determinar qué extremo del huevo será la cabeza, qué cola2) dividir el embrión a lo largo del eje anteroposterior en 17 segmentos3) dividir cada segmento en la parte delantera, media y trasera4) marque la cabeza (6 segmentos), el cofre (3 segmentos) y el abdomen (8 segmentos)5) marque las diferencias individuales de los segmentos Lared de genes que se ocupa de esto tiene un dispositivo modular. Cada módulo realiza su propio paso del algoritmo general. La mosca Drosophila resultó ser un objeto muy conveniente para estudiar las redes de desarrollo de genes, ya que tiene deformidades hereditarias muy brillantes y reveladoras. Por ejemplo, Antennapedia mutación en la que las antenas se transforman en las piernas o los VROTMNENOGIproboscipedia, cuando parte de la trompa se convierte en patas. La mutación bithorax da una mosca de cuatro alas, que, además de las alas habituales en el segundo segmento torácico, tiene un segundo par de alas en el tercer segmento torácico.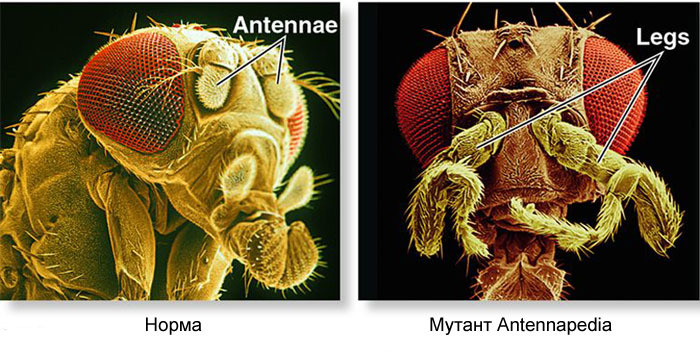
 Tales mutaciones, en las cuales una parte del cuerpo se vuelve muy similar a otra, se llaman "homeosis". Comenzando con estos mutantes, la genética finalmente descifró toda la red de genes que controlan el desarrollo de la mosca. Este trabajo ganó el Premio Nobel en 1995.Todo comienza con los genes de polaridad del huevo: bicoides y nanos. Funcionan incluso antes de la fertilización del óvulo, mientras madura en el ovario de la mosca de la madre. Las células auxiliares, que bombean nutrientes al huevo, se encuentran en un lado del huevo, por lo que el huevo tiene dos extremos diferentes desde el principio. El ARN bicoide matricial se acumula en el extremo anterior del huevo, donde está unido y no puede alejarse nadando. El ARNm de nanos también se fija en el extremo posterior. Después de la fertilización, las proteínas comienzan a sintetizarse usando estos ARNm. Sus moléculas se extienden por todo el huevo. Dado que la síntesis de proteínas bicoides y nanos se produce en un extremo del huevo y se descompone en todas partes, disminuye el gradiente de proteínas bicoides, disminuyendo desde el extremo anterior del embrión hasta el posterior, y el gradiente nanos desde el extremo posterior al anterior.
Tales mutaciones, en las cuales una parte del cuerpo se vuelve muy similar a otra, se llaman "homeosis". Comenzando con estos mutantes, la genética finalmente descifró toda la red de genes que controlan el desarrollo de la mosca. Este trabajo ganó el Premio Nobel en 1995.Todo comienza con los genes de polaridad del huevo: bicoides y nanos. Funcionan incluso antes de la fertilización del óvulo, mientras madura en el ovario de la mosca de la madre. Las células auxiliares, que bombean nutrientes al huevo, se encuentran en un lado del huevo, por lo que el huevo tiene dos extremos diferentes desde el principio. El ARN bicoide matricial se acumula en el extremo anterior del huevo, donde está unido y no puede alejarse nadando. El ARNm de nanos también se fija en el extremo posterior. Después de la fertilización, las proteínas comienzan a sintetizarse usando estos ARNm. Sus moléculas se extienden por todo el huevo. Dado que la síntesis de proteínas bicoides y nanos se produce en un extremo del huevo y se descompone en todas partes, disminuye el gradiente de proteínas bicoides, disminuyendo desde el extremo anterior del embrión hasta el posterior, y el gradiente nanos desde el extremo posterior al anterior.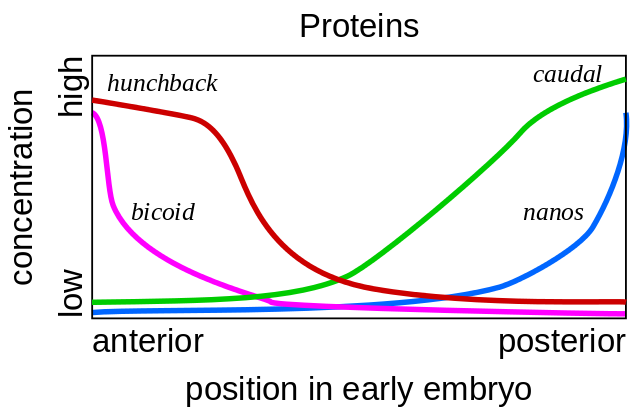 Si se introduce un ARNm bicoide en un lugar diferente del huevo con una micropipeta, allí crecerá una segunda cabeza. Si el ARNm bicoide se desactiva artificialmente, se obtiene un embrión sin cabeza y con dos asnos.Además, las proteínas bicoides y nanos incluyen los genes del embrión en sí, es decir, se comienza a leer un nuevo ARNm de ellos). Los genes gap son los primeros en comenzar a funcionar, desde el gap inglés, el gap. Los dos primeros genes de este grupo, jorobado y caudal, forman gradientes de cabeza a cola (jorobado) y de cola a cabeza (caudal). Pero sus gradientes son más suaves que los de bicoides y nanos.
Si se introduce un ARNm bicoide en un lugar diferente del huevo con una micropipeta, allí crecerá una segunda cabeza. Si el ARNm bicoide se desactiva artificialmente, se obtiene un embrión sin cabeza y con dos asnos.Además, las proteínas bicoides y nanos incluyen los genes del embrión en sí, es decir, se comienza a leer un nuevo ARNm de ellos). Los genes gap son los primeros en comenzar a funcionar, desde el gap inglés, el gap. Los dos primeros genes de este grupo, jorobado y caudal, forman gradientes de cabeza a cola (jorobado) y de cola a cabeza (caudal). Pero sus gradientes son más suaves que los de bicoides y nanos.¿Por qué los genes se llaman tan extrañamente?. . , hunchback — «», tailless — «», kruppel - — «», bithorax – « ». Sonic hedgehog , .
Los cuatro genes restantes del grupo gap forman cada uno una o dos bandas amplias (más de un segmento) con límites bastante claros. Se incorporan mediante productos proteicos de genes superiores (bicoides, nanos, jorobados y caudales). Un apagado completo de uno de los genes de este grupo da un embrión acortado, que carece de varios segmentos, aquellos en los que este gen funcionaba normalmente.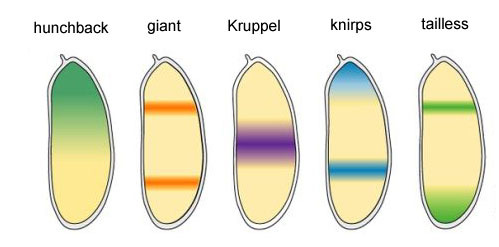 Áreas de actividad de los genes de bloqueo de huecos¿Por qué resulta que cuatro genes de bloqueo de huecos (kruppel, gigante, knirps y sin cola) dividen el embrión en bandas, en cada uno de los cuales funciona uno de los cuatro genes?En primer lugar, todos suprimen la actividad del otro. En una sola célula, una condición sería estable en la que un gen de dicho grupo trabaja activamente y suprime todos los demás (desencadenante). Pero el huevo en desarrollo es más complejo. En él, en este momento, miles de núcleos celulares individuales flotan en el líquido intracelular total (citoplasma). El tamaño del huevo es lo suficientemente grande como para que la velocidad de descomposición de las proteínas reguladoras sea comparable a su velocidad de difusión al otro extremo del huevo. Con una combinación de difusión y supresión mutua, aparece otro estado estable cuando el huevo se divide en varias bandas, en cada una de las cuales funciona un gen del grupo. Cuanto más lenta sea la difusión de proteínas o más rápido el desglose, más pequeñas serán estas rayas o manchas.Alan Turing (sí, quien inventó la "máquina de Turing") predijo la participación de tales sistemas (se les llama reacción-difusión) en el marcado del cuerpo de los animales en 1952. En ese momento, se sabía poco sobre genes y proteínas, pero las ecuaciones que describen estructuras rayadas y manchadas fueron deducidas correctamente por Turing.Aquí o aquí, puedes jugar con un modelo simple de reacción-difusión de dos sustancias y sentir cómo puede comportarse.
Áreas de actividad de los genes de bloqueo de huecos¿Por qué resulta que cuatro genes de bloqueo de huecos (kruppel, gigante, knirps y sin cola) dividen el embrión en bandas, en cada uno de los cuales funciona uno de los cuatro genes?En primer lugar, todos suprimen la actividad del otro. En una sola célula, una condición sería estable en la que un gen de dicho grupo trabaja activamente y suprime todos los demás (desencadenante). Pero el huevo en desarrollo es más complejo. En él, en este momento, miles de núcleos celulares individuales flotan en el líquido intracelular total (citoplasma). El tamaño del huevo es lo suficientemente grande como para que la velocidad de descomposición de las proteínas reguladoras sea comparable a su velocidad de difusión al otro extremo del huevo. Con una combinación de difusión y supresión mutua, aparece otro estado estable cuando el huevo se divide en varias bandas, en cada una de las cuales funciona un gen del grupo. Cuanto más lenta sea la difusión de proteínas o más rápido el desglose, más pequeñas serán estas rayas o manchas.Alan Turing (sí, quien inventó la "máquina de Turing") predijo la participación de tales sistemas (se les llama reacción-difusión) en el marcado del cuerpo de los animales en 1952. En ese momento, se sabía poco sobre genes y proteínas, pero las ecuaciones que describen estructuras rayadas y manchadas fueron deducidas correctamente por Turing.Aquí o aquí, puedes jugar con un modelo simple de reacción-difusión de dos sustancias y sentir cómo puede comportarse.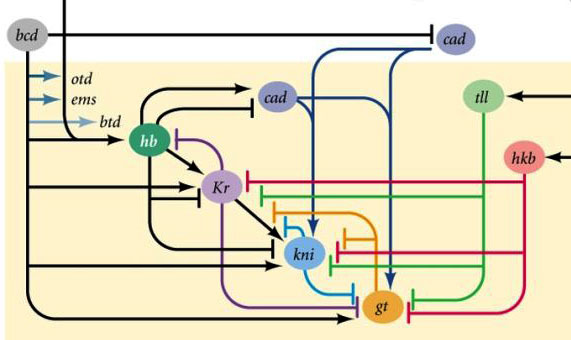 Enlaces entre genes gap y genes de polaridad del huevo. Bcd - bicoide, hb - jorobado, cad - caudal, Kr - Kruppel, kni - knirps, gt - gigante, tll - sin cola.La supresión de los genes del otro solo marcará al embrión en algunas franjas, pero la ubicación de las bandas de actividad de los cuatro genes entre sí y los extremos del embrión será muy variable. Para que las tiras estén siempre en su lugar (por ejemplo, para los nudos, una franja en la cabeza y la otra en la mitad posterior entre el kruppel y las tiras gigantes), necesitamos una segunda señal de control: gradientes de bicoides y otras proteínas de polaridad del huevo. Las funciones de entrada de los genes gap para estos reguladores tienen máximos en diferentes niveles de la señal de entrada (derivamos dicha función de entrada al final del segundo artículo ). Por lo tanto, los genes gap se activan por su nivel de proteínas bicoides, jorobadas y caudales, y sus zonas de actividad se ubican naturalmente desde la cabeza hasta la cola del embrión.Después de que los genes gap hayan realizado una marca aproximada del embrión en 7 bandas (no coinciden ni con los segmentos ni con la cabeza, el tórax y el abdomen), se activa el siguiente módulo: regla de pares ("reglas de reglas de pares"). Ya divide el embrión en segmentos, que deberían ser 17. El módulo de regla de pares consta de cinco genes, y todos, como habrás adivinado, se suprimen entre sí. Cada uno de estos genes funciona en todos los segmentos pares o impares: las proteínas codificadas por ellos se descomponen más rápido que las proteínas del grupo gap, y las tiras son correspondientemente más estrechas. Apagar parte de los genes de este grupo produce un embrión acortado, en el cual todos los segmentos pares o impares desaparecen.
Enlaces entre genes gap y genes de polaridad del huevo. Bcd - bicoide, hb - jorobado, cad - caudal, Kr - Kruppel, kni - knirps, gt - gigante, tll - sin cola.La supresión de los genes del otro solo marcará al embrión en algunas franjas, pero la ubicación de las bandas de actividad de los cuatro genes entre sí y los extremos del embrión será muy variable. Para que las tiras estén siempre en su lugar (por ejemplo, para los nudos, una franja en la cabeza y la otra en la mitad posterior entre el kruppel y las tiras gigantes), necesitamos una segunda señal de control: gradientes de bicoides y otras proteínas de polaridad del huevo. Las funciones de entrada de los genes gap para estos reguladores tienen máximos en diferentes niveles de la señal de entrada (derivamos dicha función de entrada al final del segundo artículo ). Por lo tanto, los genes gap se activan por su nivel de proteínas bicoides, jorobadas y caudales, y sus zonas de actividad se ubican naturalmente desde la cabeza hasta la cola del embrión.Después de que los genes gap hayan realizado una marca aproximada del embrión en 7 bandas (no coinciden ni con los segmentos ni con la cabeza, el tórax y el abdomen), se activa el siguiente módulo: regla de pares ("reglas de reglas de pares"). Ya divide el embrión en segmentos, que deberían ser 17. El módulo de regla de pares consta de cinco genes, y todos, como habrás adivinado, se suprimen entre sí. Cada uno de estos genes funciona en todos los segmentos pares o impares: las proteínas codificadas por ellos se descomponen más rápido que las proteínas del grupo gap, y las tiras son correspondientemente más estrechas. Apagar parte de los genes de este grupo produce un embrión acortado, en el cual todos los segmentos pares o impares desaparecen.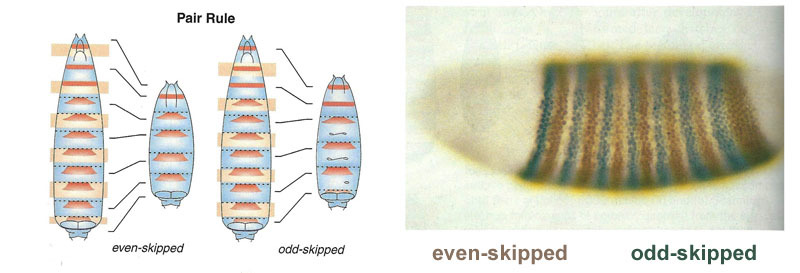 Las zonas de actividad de los genes del grupo de regla de pareja. La foto de la derecha tiene 14 tiras, no 17, porque en los primeros tres segmentos de la cabeza funciona otro sistema de marcado.El módulo de regla de pares en sí, sin la participación de gap y bicoid, puede marcar el embrión en tiras estrechas, pero el número de estas tiras sería inestable: los segmentos pueden ser 12, 18 e incluso 20. Normalmente, los genes de regla de pares son difíciles de controlar genes gap, y el primer marcado aproximado (para 7 zonas) ayuda a realizar el marcado final de manera precisa y confiable (para 14 + 3 segmentos).El siguiente punto en el algoritmo es marcar el frente, el medio y la parte posterior de cada segmento. Esto se realiza por genes de polaridad de segmento. También se suprimen entre sí y se activan mediante módulos superiores de la red de genes (regla de pares y gap). Solo se seleccionan los parámetros de difusión y descomposición de sus productos para que las tiras sean muy delgadas: medio segmento. Suprimirlos entre sí es más débil y más organizado que con los genes de regla de pareja. Por lo tanto, sus tiras de actividad se superponen parcialmente y en cada segmento se obtiene una "línea" frecuente de tiras superpuestas de manera diferente.La desactivación de los genes de polaridad de segmento generalmente no conduce al acortamiento del embrión y la pérdida de sus partes (como es típico para la brecha y la regla de pares), sino a efectos más sutiles. Por lo general, en este caso, la mitad de cada segmento en el que se suponía que este gen debía funcionar se convierte en una imagen especular de la otra mitad. Esto es claramente visible, porque la larva de la mosca tiene hileras de pequeñas espinas dirigidas hacia atrás, para la conveniencia de gatear. Los mutantes en el segmento de los genes de polaridad tienen filas de espinas dirigidas hacia adelante.Finalmente, es el turno de los genes más famosos y descubiertos por primera vez de esta red: el módulo hox. Su trabajo le da a los segmentos individualidad para que las antenas y la trompa crezcan en los segmentos de la cabeza, y las piernas y las alas crezcan en los segmentos del pecho. Los genes hox, a diferencia de los módulos anteriores, no inhiben la actividad de los demás y pueden trabajar juntos en el mismo segmento embrionario. Una extraña propiedad de los genes hox que aún no se ha explicado: están ubicados en el cromosoma en un grupo apretado y en el mismo orden en que trabajan en el cuerpo del embrión.
Las zonas de actividad de los genes del grupo de regla de pareja. La foto de la derecha tiene 14 tiras, no 17, porque en los primeros tres segmentos de la cabeza funciona otro sistema de marcado.El módulo de regla de pares en sí, sin la participación de gap y bicoid, puede marcar el embrión en tiras estrechas, pero el número de estas tiras sería inestable: los segmentos pueden ser 12, 18 e incluso 20. Normalmente, los genes de regla de pares son difíciles de controlar genes gap, y el primer marcado aproximado (para 7 zonas) ayuda a realizar el marcado final de manera precisa y confiable (para 14 + 3 segmentos).El siguiente punto en el algoritmo es marcar el frente, el medio y la parte posterior de cada segmento. Esto se realiza por genes de polaridad de segmento. También se suprimen entre sí y se activan mediante módulos superiores de la red de genes (regla de pares y gap). Solo se seleccionan los parámetros de difusión y descomposición de sus productos para que las tiras sean muy delgadas: medio segmento. Suprimirlos entre sí es más débil y más organizado que con los genes de regla de pareja. Por lo tanto, sus tiras de actividad se superponen parcialmente y en cada segmento se obtiene una "línea" frecuente de tiras superpuestas de manera diferente.La desactivación de los genes de polaridad de segmento generalmente no conduce al acortamiento del embrión y la pérdida de sus partes (como es típico para la brecha y la regla de pares), sino a efectos más sutiles. Por lo general, en este caso, la mitad de cada segmento en el que se suponía que este gen debía funcionar se convierte en una imagen especular de la otra mitad. Esto es claramente visible, porque la larva de la mosca tiene hileras de pequeñas espinas dirigidas hacia atrás, para la conveniencia de gatear. Los mutantes en el segmento de los genes de polaridad tienen filas de espinas dirigidas hacia adelante.Finalmente, es el turno de los genes más famosos y descubiertos por primera vez de esta red: el módulo hox. Su trabajo le da a los segmentos individualidad para que las antenas y la trompa crezcan en los segmentos de la cabeza, y las piernas y las alas crezcan en los segmentos del pecho. Los genes hox, a diferencia de los módulos anteriores, no inhiben la actividad de los demás y pueden trabajar juntos en el mismo segmento embrionario. Una extraña propiedad de los genes hox que aún no se ha explicado: están ubicados en el cromosoma en un grupo apretado y en el mismo orden en que trabajan en el cuerpo del embrión.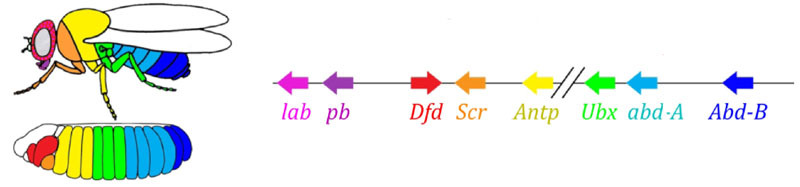 El marcado del embrión se basa en el principio "cuanto más lejos del tórax, más genes hox y más activo es": la actividad del gen abdominal-A comienza desde el segundo segmento abdominal, crece a 4 y continúa al nivel máximo en todos los segmentos posteriores; El gen abdominal-B funciona desde el 5º hasta el 8º segmento del abdomen, aumentando la actividad en cada segmento posterior. El gen ultrabithorax está incluido en la fuerza media en el tercer segmento torácico y a plena capacidad en todo el abdomen. Cuando está desactivado en mutantes de bithorax, las células del segmento 3 torácico no saben que son el segmento 3 torácico, el mismo conjunto de genes hox funciona en ellos que en el 2 torácico (es decir, solo antenapedia) y el desarrollo del segmento 3 torácico continúa programa 2 cofres: con alas en lugar de escarabajos de tierra y poderosos músculos voladores.Si desactiva todos los genes hox en un segmento separado, resultará feo, pero sobre todo se verá como 1 cofre, con patas y sin alas. Esto se debe a que los ancestros distantes de los insectos eran como trilobites o milpiés. Tenían muchos segmentos idénticos con patas. Posteriormente, su cuerpo se acortó, la parte posterior perdió piernas, convirtiéndose en el abdomen, y varios segmentos frontales se unieron a la cabeza y sus piernas se convirtieron en partes del aparato oral (mandíbulas, maxilares y labio inferior).
El marcado del embrión se basa en el principio "cuanto más lejos del tórax, más genes hox y más activo es": la actividad del gen abdominal-A comienza desde el segundo segmento abdominal, crece a 4 y continúa al nivel máximo en todos los segmentos posteriores; El gen abdominal-B funciona desde el 5º hasta el 8º segmento del abdomen, aumentando la actividad en cada segmento posterior. El gen ultrabithorax está incluido en la fuerza media en el tercer segmento torácico y a plena capacidad en todo el abdomen. Cuando está desactivado en mutantes de bithorax, las células del segmento 3 torácico no saben que son el segmento 3 torácico, el mismo conjunto de genes hox funciona en ellos que en el 2 torácico (es decir, solo antenapedia) y el desarrollo del segmento 3 torácico continúa programa 2 cofres: con alas en lugar de escarabajos de tierra y poderosos músculos voladores.Si desactiva todos los genes hox en un segmento separado, resultará feo, pero sobre todo se verá como 1 cofre, con patas y sin alas. Esto se debe a que los ancestros distantes de los insectos eran como trilobites o milpiés. Tenían muchos segmentos idénticos con patas. Posteriormente, su cuerpo se acortó, la parte posterior perdió piernas, convirtiéndose en el abdomen, y varios segmentos frontales se unieron a la cabeza y sus piernas se convirtieron en partes del aparato oral (mandíbulas, maxilares y labio inferior).Modelos matemáticos de la red genética para el marcado de embriones.
En el caso del operón de lactosa y los bucles de enlace directo en Escherichia coli, es posible construir modelos matemáticos bastante simples con un número moderado de parámetros que predicen el comportamiento de los genes para cualquier señal de entrada. Para un operón de lactosa, es suficiente:- cuatro constantes de unión (represor y activador con ADN, lactosa con represor, cAMP con activador)- cuatro parámetros de estequiometría (4 lactosa por 1 represor, 2 activador cAMP 1, 1 represor y activador por operón)- dos parámetros de regulación (el efecto del represor y el activador sobre la probabilidad de aterrizaje de la ARN polimerasa)- el nivel básico de actividad del operón (cuando no hay represor o activador)- retraso transcripcionalTotal de 12 parámetros.Veamos ahora las regiones reguladoras de, digamos, los genes Hox. En el diagrama a continuación, uno de los bloques reguladores del gen abdominal-B. Las barras coloreadas indican sitios reguladores de unión a proteínas. Todo este bloque tiene un tamaño de aproximadamente 1700 nucleótidos, 10 veces más largo que la región reguladora del operón lactosa. Tiene 39 sitios de siembra para 7 proteínas reguladoras diferentes de todos los bloques superiores de esta red de genes. Y abdominal-B tiene cuatro de estos bloques reguladores. Se encienden secuencialmente del 5 ° al 8 ° segmento del abdomen y crean un aumento gradual en el nivel de proteína abdominal-B en estos segmentos. Es decir, solo para el modelo de este gen, en el buen sentido, necesita más de 100 constantes de unión, el mismo número de parámetros de regulación y un número desconocido de parámetros para describir la interacción entre diferentes entradas, en el peor de los casos, hasta 1,500 interacciones por pares entre todos los sitios de desembarque de proteínas en uno unidad reguladora.Es decir, el modelo completo de esta red de genes ciertamente tendrá más de mil parámetros. Y este es el problema principal: no hay tantas señales de salida que podamos medir en el experimento; de hecho, solo los niveles de cada una de las 25 proteínas y su distribución a lo largo del embrión. Por lo tanto, nadie intentó simular esta red con la misma granularidad que el operón lactosa.Puede intentar simplificar la tarea y describir el efecto de cada proteína reguladora en el gen con solo tres parámetros (enlace, estequiometría y parámetro de regulación). Puede seleccionar estos parámetros (por ejemplo, utilizando cadenas de Markov) para que el modelo reproduzca con precisión las franjas reales de actividad genética. Para los primeros dos bloques de la red (genes de polaridad del huevo y genes gap), tales modelos incluso se hacen ( por ejemplo ).El problema es que los diferentes inicios del algoritmo de selección dan diferentes parámetros del modelo, ¡pero todos funcionan correctamente y reproducen la actividad de los genes en un brote de mosca real! Estos modelos tienen características comunes (por ejemplo, todos los genes gap se suprimen entre sí y son activados por las proteínas madre), pero no hay menos diferencias que similitudes.Y dado que los diferentes intentos de modelado dan resultados diferentes, es poco probable que al menos uno de ellos coincida con la realidad. Peor aún, tales modelos reproducen el funcionamiento normal, pero pierden mucho en predecir los resultados de las mutaciones.En la siguiente parte, hablaremos sobre por qué estos problemas con el estudio de las redes de genes son inevitables en biología (¡y la red de marcado de embriones de Drosophila es una de las más simples de estudiar!), ¿Qué se puede hacer al respecto y qué ingenieros deben aprender de los biólogos?
Las barras coloreadas indican sitios reguladores de unión a proteínas. Todo este bloque tiene un tamaño de aproximadamente 1700 nucleótidos, 10 veces más largo que la región reguladora del operón lactosa. Tiene 39 sitios de siembra para 7 proteínas reguladoras diferentes de todos los bloques superiores de esta red de genes. Y abdominal-B tiene cuatro de estos bloques reguladores. Se encienden secuencialmente del 5 ° al 8 ° segmento del abdomen y crean un aumento gradual en el nivel de proteína abdominal-B en estos segmentos. Es decir, solo para el modelo de este gen, en el buen sentido, necesita más de 100 constantes de unión, el mismo número de parámetros de regulación y un número desconocido de parámetros para describir la interacción entre diferentes entradas, en el peor de los casos, hasta 1,500 interacciones por pares entre todos los sitios de desembarque de proteínas en uno unidad reguladora.Es decir, el modelo completo de esta red de genes ciertamente tendrá más de mil parámetros. Y este es el problema principal: no hay tantas señales de salida que podamos medir en el experimento; de hecho, solo los niveles de cada una de las 25 proteínas y su distribución a lo largo del embrión. Por lo tanto, nadie intentó simular esta red con la misma granularidad que el operón lactosa.Puede intentar simplificar la tarea y describir el efecto de cada proteína reguladora en el gen con solo tres parámetros (enlace, estequiometría y parámetro de regulación). Puede seleccionar estos parámetros (por ejemplo, utilizando cadenas de Markov) para que el modelo reproduzca con precisión las franjas reales de actividad genética. Para los primeros dos bloques de la red (genes de polaridad del huevo y genes gap), tales modelos incluso se hacen ( por ejemplo ).El problema es que los diferentes inicios del algoritmo de selección dan diferentes parámetros del modelo, ¡pero todos funcionan correctamente y reproducen la actividad de los genes en un brote de mosca real! Estos modelos tienen características comunes (por ejemplo, todos los genes gap se suprimen entre sí y son activados por las proteínas madre), pero no hay menos diferencias que similitudes.Y dado que los diferentes intentos de modelado dan resultados diferentes, es poco probable que al menos uno de ellos coincida con la realidad. Peor aún, tales modelos reproducen el funcionamiento normal, pero pierden mucho en predecir los resultados de las mutaciones.En la siguiente parte, hablaremos sobre por qué estos problemas con el estudio de las redes de genes son inevitables en biología (¡y la red de marcado de embriones de Drosophila es una de las más simples de estudiar!), ¿Qué se puede hacer al respecto y qué ingenieros deben aprender de los biólogos?