En mi publicación anterior , se consideraron dos preguntas: los fundamentos de la biología molecular y los métodos para crear las construcciones que necesitamos que se basan en vectores plasmídicos. Ahora necesita descubrir cómo introducir un vector plasmídico en una célula bacteriana, es decir, para hacer una "transformación". Al mismo tiempo, aprenderemos algo sobre la estructura de la membrana bacteriana y cómo se puede superar, así como por qué algunas bacterias se llaman competentes, cómo todo esto se asocia con toxinas bacterianas y resistencia bacteriana a los antibióticos, y muchos otros hechos interesantes.
Entonces, tenemos un vector plasmídico en nuestras manos y hay una cepa bacteriana adecuada para este vector, queda por introducir el vector en la bacteria.
En un
artículo anterior , se mencionó el
trabajo de Frederick Griffith , que demuestra claramente el hecho de que el ADN puede penetrar espontáneamente en las células bacterianas. La capacidad de una célula bacteriana para absorber una molécula de ADN ambiental se llama "competencia" (lo que genera muchas bromas sobre el hecho de que incluso los in vitro unicelulares son competentes, y su colega de laboratorio no lo es). Si la absorción no ocurre bajo la influencia de algunas condiciones creadas artificialmente, entonces estamos hablando de "competencia natural". ¿Cómo se produce un transporte de ADN similar dentro de una bacteria?
1) La estructura de la membrana celular de las bacterias.
Para empezar, consideramos brevemente la estructura de la membrana de las bacterias grampositivas y gramnegativas.
El caparazón de una bacteria gramnegativa consta de dos bicapas lipídicas: la membrana externa y la membrana interna (la interna también se llama plasma). Entre ellos se encuentra el espacio periplásmico lleno de proteínas y peptidoglicanos que realizan funciones importantes.
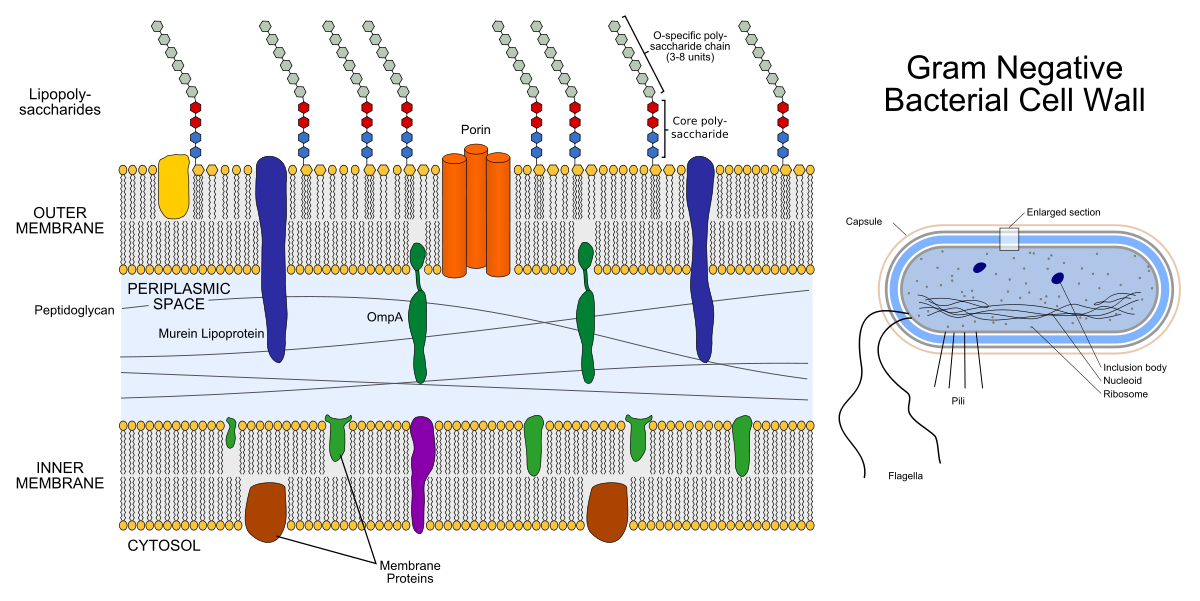 A la izquierda está la estructura de la membrana de la bacteria gramnegativa. Los artilugios coloreados dentro de las membranas son proteínas de membrana, y las cadenas que sobresalen son lipopolisacáridos. A la derecha está la estructura de la bacteria gramnegativa misma.
A la izquierda está la estructura de la membrana de la bacteria gramnegativa. Los artilugios coloreados dentro de las membranas son proteínas de membrana, y las cadenas que sobresalen son lipopolisacáridos. A la derecha está la estructura de la bacteria gramnegativa misma.El peptidoglucano es un heteropolímero que consiste en N-acetilglucosamina reticulada covalentemente (NAG) y ácido N-acetilmurámico (NAM). Resulta que la cadena ...- NAG-NAM-NAG-NAM-NAG- .... "Hetero" en la palabra "heteropolímero" significa que el polímero no consta de los mismos elementos (como el polietileno), sino de dos elementos diferentes. La enzima sintetizadora de peptidoglucano, la transpeptidasa, es un objetivo para los antibióticos β-lactámicos (antibióticos de penicilina y cefalosporinas): inhiben su actividad, como resultado del proceso de división bacteriana, no hay suficiente peptidoglucano para ambas células hijas. La capa de peptidoglucano también es un objetivo para la enzima lisozima: destruye el enlace covalente entre NAG y NAM. En medicina, la lisozima se usa como antiséptico, por ejemplo, es un componente activo de algunos medicamentos para el tratamiento del dolor de garganta, y en la industria alimentaria puede usarse como conservante (suplemento alimenticio E1105, esto es lisozima).

La envoltura de las bacterias grampositivas consta solo de una membrana interna (plasma), un espacio periplásmico muy delgado y una capa externa de peptidoglucano más gruesa que las bacterias gramnegativas.
Tales obstáculos que se interponen en el camino de cualquier compuesto en el citoplasma bacteriano son insuperables para todo, excepto el agua, así como para las pequeñas moléculas no polares e hidrófobas (oxígeno molecular y nitrógeno, dióxido de carbono y otros). La penetración de estos compuestos a través de la cubierta se llama "transporte pasivo" ("difusión simple"), siempre ocurre a lo largo del gradiente de concentración. Las moléculas no polares pueden pasar libremente a través de las membranas porque el volumen interno de las membranas también es no polar (dentro de la membrana están formadas por residuos hidrófobos de ácidos grasos), y los compuestos no polares se disuelven bien en solventes no polares (alcohol, acetona, ácidos grasos).
Una imagen cómica en la que se juega la hidrofobicidad (hidrofobia) de los residuos de ácidos grasos dentro de una membrana de dos capas.A su vez, las moléculas polares grandes (aminoácidos, monosacáridos, nucleótidos), partículas cargadas (iones) y macromoléculas (ADN, proteínas) no pueden penetrar a través de la membrana celular así, ya que son muy solubles solo en solventes polares (por ejemplo, en agua) . Entonces, ¿cómo entran las moléculas de ADN dentro de una célula bacteriana?
2) Mecanismos de transporte inducido de compuestos a las células desde el medio ambiente.
En primer lugar, tenemos en nuestras manos el hecho obvio de que la bacteria no puede vivir solo del agua, el oxígeno y el dióxido de carbono, para la vida necesita muchos otros compuestos. Por lo tanto, hay formas de administrar estos compuestos a la célula. Dichos métodos son "difusión facilitada" y "transporte activo".
2.1) Difusión de luz
Un ejemplo de difusión facilitada (aunque este caso particular, por el contrario, es perjudicial para las bacterias) es el transporte de iones de potasio a través de la membrana de una célula bacteriana en combinación con la valinomicina, un antibiótico que es sintetizado por algunos hongos. La valinomicina libre no puede penetrar en la célula, ya que los grupos polares están expuestos externamente. El ion potasio también es un "camino cerrado".
 Una escultura que representa la estructura tridimensional de un complejo de valinomicina e iones de potasio (el ión se representa como una bola en el centro). Se instaló frente a la entrada de M. M. Shemyakin y Yu. A. Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences (Moscú), en el que se estableció el mecanismo de la actividad antibacteriana de la valinomicina y la estructura tridimensional de su complejo con iones de potasio.
Una escultura que representa la estructura tridimensional de un complejo de valinomicina e iones de potasio (el ión se representa como una bola en el centro). Se instaló frente a la entrada de M. M. Shemyakin y Yu. A. Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences (Moscú), en el que se estableció el mecanismo de la actividad antibacteriana de la valinomicina y la estructura tridimensional de su complejo con iones de potasio.Pero después de la formación de un complejo de valinomicina con un ion de potasio, cambia su estructura tridimensional para que toda su superficie externa se vuelva hidrófoba (no polar) y todos los grupos hidrofílicos se oculten en su interior. En este caso, el ión potasio está oculto dentro de la molécula de valinomicina; por lo tanto, el complejo de valinomicina + ión potasio puede penetrar a través de la membrana. El complejo puede descomponerse en valinomicina e iones libres en cualquier momento, tanto dentro como fuera de la célula, pero generalmente la difusión de iones facilitada conduce a la igualación de su concentración en los espacios intracelulares y extracelulares, y la célula controla el transporte de iones por una razón: su alta concentración fatal En esto se basa el efecto antibacteriano de la valinomicina.
Representación esquemática del antibiótico fúngico valinomicina complejado con iones de potasio.Otro ejemplo son los poros no controlados, que son proteínas transmembrana grandes que permiten que ciertas sustancias pasen libremente a través de la membrana según el gradiente. Un ejemplo de tal poro transmembrana es el
Staphylococcus aureus Staphylococcus aureus α-toxina (otro nombre para la toxina: α-hemolisina. Consiste en las partes "hemo" y "lisina". "Hemo" significa glóbulos rojos; "lisina" significa de la palabra "lisis", es decir, "muerte celular")
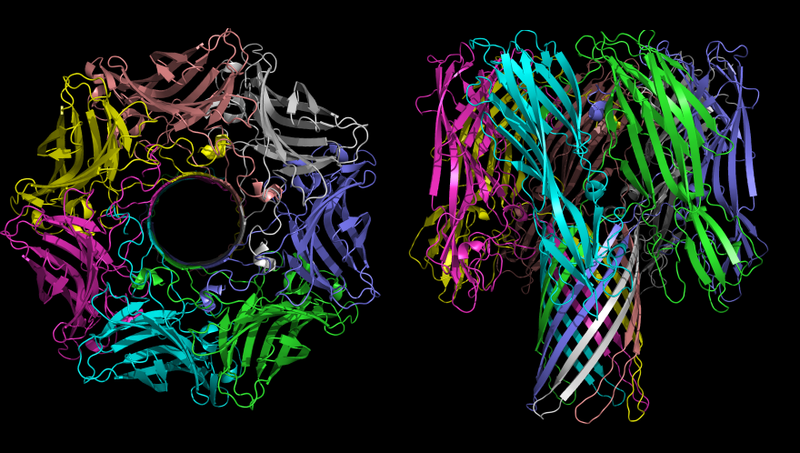 La estructura tridimensional del oligómero de α-hemolisina, la toxina de la bacteria Staphylococcus aureus. En la imagen de la izquierda, el tiempo es claramente visible.
La estructura tridimensional del oligómero de α-hemolisina, la toxina de la bacteria Staphylococcus aureus. En la imagen de la izquierda, el tiempo es claramente visible.La función principal de la α-hemolisina en el proceso patogénico es la creación de canales no controlados en la membrana celular, que, según las condiciones ambientales, pueden pasar iones monovalentes, iones de calcio o ATP. Como se mencionó anteriormente, el desequilibrio iónico es extremadamente tóxico, y el transporte de ATP
desde la célula es dañino, ya que el ATP es un enlace indispensable en casi todos los procesos químicos de cualquier célula.
2.2) Transporte activo
Como se puede ver en los ejemplos anteriores, la difusión facilitada no requiere energía. Por el contrario, el transporte activo es la absorción o liberación de algo por una célula con el gasto de energía para cada acto de transferencia. Este puede ser el transporte de grandes moléculas polares o iones, y el transporte de iones puede llevarse a cabo tanto a lo largo del gradiente de concentración como contra él. Por supuesto, el transporte activo es un proceso controlado en el que interviene un conjunto completo de proteínas intracelulares y transmembrana. Por lo tanto, primero la célula necesita gastar recursos en la síntesis de todas estas proteínas y luego gastar energía nuevamente en el uso de este sistema de transporte.
Un ejemplo de transporte activo es el sistema de transporte dependiente de energía de compuestos hidrofóbicos y anfifílicos desde una célula de regreso al medio ambiente (eflujo).
El sistema de transporte activo de un antibiótico desde una célula bacteriana hacia el exterior, que consta de tres proteínas que forman un único complejo transmembrana. La figura refleja el hecho de que la proteína transportadora de eflujo (violeta) puede "atrapar" moléculas de fármaco directamente en el citoplasma y en la membrana.Este sistema es de particular interés porque causa la resistencia múltiple de algunas bacterias a los antibióticos: la célula los elimina antes de que puedan dañarlo seriamente. Además, más de la mitad de los casos de resistencia a múltiples fármacos de
Pseudomonas aeruginosa Pseduomonas aeruginosa están asociados con el transporte activo de antibióticos. Más importante aún, ya se han descubierto sistemas de eflujo activo de "nuevos" antibióticos que se usan contra bacterias que son resistentes a los fármacos antibacterianos de uso común. Por ejemplo, el medicamento
Linezolid utilizado en el caso de la ineficacia de los antibióticos clásicos es arrojado por el sistema de flujo de salida basado en las proteínas AcrAB y TolC. Otro hecho interesante es que una bacteria inteligente no desperdicia energía en la síntesis a gran escala de proteínas del sistema de eflujo en aquellas situaciones en que no son necesarias. Pero tan pronto como aparecen las sustancias nocivas para la célula, la síntesis se activa rápidamente y la bacteria comienza a resistir activamente. Por cierto, el flujo de salida no es el único mecanismo para proteger las bacterias de los antibióticos, también escribiré un artículo sobre otros métodos.
La estructura del linezolid es un antibiótico utilizado contra cepas resistentes de bacterias grampositivas.3) Competencia de las células bacterianas.
Entonces, examinamos los mecanismos de transporte transmembrana de compuestos que no pueden penetrar esta barrera sin ayuda externa. En condiciones naturales, el transporte de ADN hacia la bacteria se lleva a cabo solo a través del transporte activo debido a la cooperación de un espectro completo de proteínas intracelulares y transmembrana. La capacidad no inducida de una célula para absorber el ADN del exterior se denomina "competencia natural".
3.1) Competencia natural de las células bacterianas.
La hipótesis más confirmada de la aparición de la competencia natural de las bacterias es la suposición de que este mecanismo permite que las bacterias sobrevivan en condiciones extremas (las condiciones extremas para una célula se denominan "estrés"). El hecho es que el estrés a menudo conduce al daño del ADN "cromosómico" de una bacteria, y si una célula quiere sobrevivir, entonces necesita de alguna manera "reparar" su "cromosoma" (dicha "reparación" se llama "reparación"). Además, se sabe que si se introduce una secuencia de ADN en la célula que es similar a alguna parte del ADN de la célula, es muy probable que integre esta "nueva" pieza en su genoma. ¿Qué pasa bajo condiciones estresantes? Algunas células mueren, convirtiéndose en donantes de ADN para aquellos de sus hermanos que aún luchan por la vida. De hecho, dentro del mismo cultivo bacteriano, es muy probable que los genomas de los representantes sean muy similares, por lo que las bacterias sobrevivientes pueden usar el ADN de los parientes muertos para reparar su genoma.
Además, bajo condiciones cómodas en el cultivo celular, solo una fracción de ellas tiene competencia natural, y la proporción de células competentes de su número total es la más grande entonces, entonces el cultivo está en una cierta fase de crecimiento: la fase
logarítmica .
 Una trama típica de bacterias en cultivo a lo largo del tiempo. La fase de registro es la fase de crecimiento, seguida de una meseta y la etapa de extinción, en la que las células ya carecen de nutrientes y se acumulan toxinas.
Una trama típica de bacterias en cultivo a lo largo del tiempo. La fase de registro es la fase de crecimiento, seguida de una meseta y la etapa de extinción, en la que las células ya carecen de nutrientes y se acumulan toxinas.Por supuesto, al investigador le gustaría hacer que el proceso de transferencia del vector plasmídico a la célula sea lo más eficiente posible: cuanto más eficiente sea el proceso de transformación, se necesitará menos vector plasmídico para garantizar buenos resultados. Por lo tanto, en condiciones de laboratorio, generalmente no dependen de la célula en sí, sino que crean células con competencia artificial.
3.2) Competencia artificial de las células bacterianas.
Existen dos métodos principales para crear la competencia artificial de las bacterias: el tratamiento de un cultivo celular con iones divalentes seguido de calentamiento a corto plazo y la llamada electroporación.
3.2.1) Tratamiento del cultivo celular con iones divalentes, seguido de calentamiento a corto plazo.
Como regla general, en el laboratorio hay un refrigerador con una temperatura de aproximadamente -80 grados centígrados (a veces se los llama "kelvinators" en honor a una de las empresas manufactureras), obstruido con tubos de ensayo con células en la fase
logarítmica . Esto se hace para no perder el tiempo creándolos cada vez que se necesitan (y lleva mucho tiempo, no funcionará más rápido que en un día). Y para que durante el almacenamiento las células no mueran debido a la congelación, se agregan crioprotectores al medio: glicerol, propilenglicol o dimetilsulfóxido.
Refrigerador de laboratorio a la vista del autor de la imagen.Cuando un investigador necesita transformar una cepa específica, elimina las células del Kelvinator y coloca el tubo en hielo. Luego, los cationes divalentes y el vector plasmídico que necesitamos se agregan a las células en la fase
logarítmica del crecimiento. El tubo de ensayo no debe dejar el hielo. Un donante típico de tales cationes es el cloruro de calcio (CaCl2), en este caso, nuestro catión divalente es Ca2 +. No hay una explicación inequívoca de cómo el Ca2 + ayuda al ADN a penetrar la bacteria:
- Algunas fuentes afirman que las altas concentraciones de Ca2 + causan lagunas en la membrana celular y el daño del ADN, y anteriormente descubrimos que el daño del ADN es una señal para que la célula se vuelva competente;
- Según otras fuentes, estos iones sirven como un "puente" que une el vector plasmídico a la célula. El hecho es que los iones con carga positiva se unen simultáneamente a grupos de polisacáridos con carga negativa en la membrana externa de las bacterias y a grupos de ADN plasmídico con carga negativa. Si no hubiera iones en el medio, entonces el ADN sería repelido desde la célula, y esto es completamente inútil para el investigador.
Es posible que ambas hipótesis sean ciertas.
El supuesto mecanismo de participación de iones Ca2 + en la transformación. Primero, los iones se unen a grupos de ADN con carga negativa (círculos amarillos) y polisacáridos anclados en la membrana (círculos amarillos). Luego, debido al ion, se forma una especie de puente: "polisacárido (-)" - "Ca ion (2+)" - "ADN (-)", que fija el ADN en la superficie celular.Después de la incubación bacteriana a una temperatura cercana a cero, se produce el llamado choque térmico: calentamiento a corto plazo del cultivo a 42 grados. Por lo general, la duración del tratamiento térmico es de 30 a 45 segundos, después de lo cual el tubo de ensayo con bacterias regresa al baño de hielo (con un choque térmico más prolongado, las células pueden morir). Las posibles razones para la efectividad del choque térmico son los mismos mecanismos: la alta temperatura puede dañar el ADN y las proteínas (esto induce competencia), y también puede crear "agujeros" en la membrana celular a través de los cuales penetran los vectores plasmídicos unidos a la membrana a través de iones de calcio.
Luego, las células se colocan en placas de Petri con medio selectivo y crecen colonias en ellas.
3.2.2) Electroporación
La base del método de
electroporación es el hecho de que el campo eléctrico puede dañar la membrana celular. Es decir, literalmente, la electroporación es la creación de poros por un campo eléctrico.
Primero, las bacterias y un vector plasmídico se colocan en un tubo especial (cubeta). Luego, la cubeta se coloca en un dispositivo especial, que por una fracción de segundo crea una diferencia de potencial de 0.1-10 kV, lo que a su vez conduce a daños reversibles en las membranas y los vectores pueden penetrar libremente en las células. Después de algún tiempo, el cultivo bacteriano se siembra en placas de Petri con medio selectivo.
Un requisito importante para el medio líquido en el que se encuentran las células y el vector en el momento de la electroporación es una baja concentración de iones: de lo contrario, puede producirse un cortocircuito.
Epílogo
Cada colonia en un plato es el descendiente proliferante de
una sola bacteria (es decir, si vemos 100 colonias en un plato, significa que inicialmente había 100 bacterias viables, que luego comenzaron a dividirse).
Después de que las colonias aparecen en los platos, el investigador selecciona una de las colonias y transfiere las células de la colonia a un nuevo medio nutritivo líquido selectivo. Después de que un nuevo cultivo celular "crece", se planta nuevamente en una nueva copa con el mismo medio nutritivo selectivo. Como resultado, obtenemos una copa en la que crecen solo los descendientes de bacterias de una sola colonia (es decir, de hecho, todos son clones de una sola bacteria). Y solo estas células se utilizarán en el futuro para obtener la proteína que necesitamos en grandes volúmenes de medio nutritivo selectivo líquido.Bueno, resumamos los resultados intermedios:- Sintetizamos el gen que necesitamos;
- Insértelo en un vector plasmídico adecuado;
- Se seleccionó y transformó una cepa productora adecuada;
- Tengo una cultura clon;
- Finalmente, transferimos parte de estos clones a un gran volumen de medio nutriente para obtener la proteína que necesitamos en un gran volumen.
El siguiente paso: aislar la proteína de la biomasa resultante en la forma más pura posible. Pero esta es una historia completamente diferente.