Automatisation à partir de l'ADN et des protéines: qu'est-ce qu'elle a en commun avec l'électronique?
(réponse du biologiste à l'article "Vous n'écrivez pas correctement les animaux" )Il y a des milliers de gènes dans chaque cellule. Ce n'est jamais qu'ils travaillent tous simultanément. Les cellules musculaires, hépatiques et cérébrales portent le même ensemble de gènes. Ils sont si différents parce que différents gènes y travaillent. Toutes les cellules peuvent activer et désactiver des gènes individuels en réponse à diverses influences externes. Autrement dit, le système qui contrôle l'activité des gènes est un tel système de contrôle automatique. Les scientifiques veulent comprendre comment fonctionne une telle automatisation afin de pouvoir la réparer et la casser. Par exemple, dans les cellules humaines, il existe un système d'environ trois cents gènes et protéines qui contrôle la division cellulaire. Quand il se décompose et que la cellule commence à se diviser tout le temps, le cancer survient.Un schéma simplifié du réseau de gènes qui régule la division des cellules humainesComment est l'automatisation intracellulaire?
Dans le «fer», on utilise généralement des impulsions électriques. Les signaux en biologie sont généralement chimiques, c'est-à-dire des changements dans la concentration de toute substance. Il y a des signaux électriques dans le système nerveux, mais ce n'est qu'un appareil pour une transmission rapide à longue distance, semblable à la fibre optique dans la technologie. La transmission d'une impulsion nerveuse d'une cellule à l'autre se produit sous forme chimique, la mémoire à long terme et l'intégration complexe des signaux dans un neurone est également chimique.Là où il y a des signaux, il y aura des fils - les signaux ne sont pas à leur place et le bruit est une ordure aléatoire collant aux signaux. En électronique, l'arrivée des signaux à l'adresse est assurée par des fils et une isolation. La diaphonie se produit en raison du couplage capacitif et inductif parasite entre les conducteurs. Dans une cellule bactérienne, son contenu est plus ou moins uniformément mélangé, et tout signal chimique (et il y en a des milliers) est disponible en tout point. Les cellules des animaux et des plantes sont divisées en compartiments de composition chimique différente, mais les types de ces compartiments sont inférieurs à une douzaine et contiennent chacun des centaines et des milliers de substances différentes.L'arrivée du signal au bon endroit dans la cellule est due à la reconnaissance moléculaire sur le principe du "key-lock". Une molécule de protéine peut reconnaître une autre protéine, une protéine peut reconnaître une séquence d'ADN spécifique, une protéine peut reconnaître de petites molécules telles que les sucres. Cette reconnaissance n'est pas toujours parfaitement précise, donc des molécules de forme similaire à un partenaire de reconnaissance régulier créeront des astuces. En plus des interférences, il y a des bruits dans l'électronique. Ils surviennent en raison du mouvement thermique des atomes, qui introduit une fraction de chaos dans le mouvement des électrons. Dans une cellule, le bruit chimique est également associé au mouvement thermique. Lorsque le signal est faible, par exemple, dix morceaux de molécules de ce type sur toute la cellule, ces molécules seront dispersées de manière aléatoire dans la cellule, et non le fait de manière uniforme. En conséquence, leur concentration dans un coin de la cellule fluctuera de manière aléatoire, et ce sera du bruit.Qu'est-ce qui est régulé exactement dans une cellule vivante?
Une cage peut être considérée comme une usine qui produit tout l'équipement dont il a besoin et peut en assembler une seconde. L'équipement principal est les ribosomes - des machines CNC qui collectent toutes les protéines. La structure de la protéine collectée est enregistrée sur un ARN matriciel (ARNm). Le ribosome se déplace le long de l'ARNm et, selon ses instructions, recueille une chaîne d'une nouvelle protéine à partir d'unités individuelles - les acides aminés. La chaîne protéique émergeant du ribosome est pliée en une boule compacte de protéine mature, qui commence à fonctionner. Les protéines peuvent accélérer les réactions chimiques (enzymes), ainsi que le transport de substances, la signalisation, la défense et bien d'autres choses.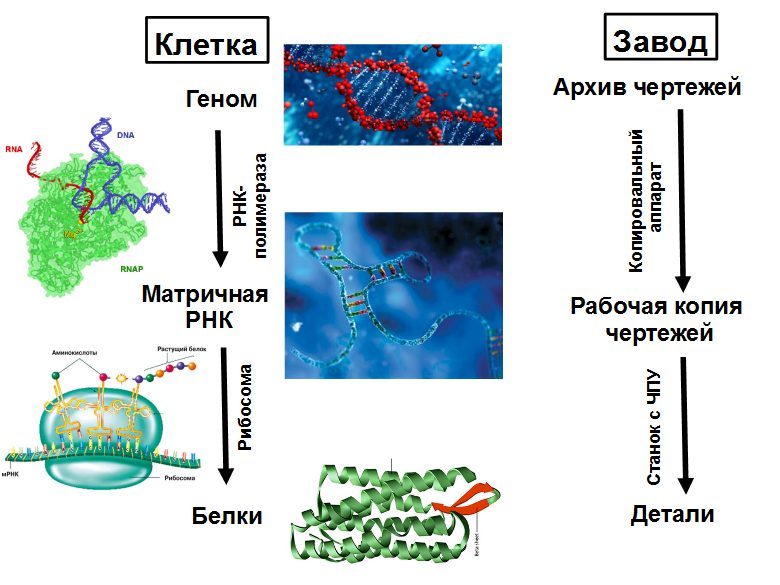
Éléments logiques sur l'ADN et les protéines
L'exemple le plus étudié de changement de gène est peut-être l'opéron lactose d'Escherichia coli. Le français Jacob et Mono pour ses recherches ont reçu le prix Nobel en 1965. E. coli, comme son nom l'indique, vit dans l'intestin. Il peut s'agir des intestins d'une grande variété d'animaux, des abeilles aux humains. Là, elle doit manger ce que le propriétaire a obtenu, afin qu'elle puisse manger de nombreux nutriments différents.E. coli au microscope et sur boîte de Pétri.E. coli peut se développer, par exemple, sur un milieu nutritif composé uniquement de glucose et de sels minéraux et produire tous les acides aminés et vitamines dont il a besoin. Au lieu du glucose, il peut y avoir d'autres sucres (fructose, sucre de malt - maltose, sucre de lait - lactose et une douzaine d'autres), ainsi que des polysaccharides, comme l'amidon.Pour absorber chacun de ces sucres, vous avez besoin de vos propres enzymes. La production de ces enzymes ne devrait commencer que lorsque le sucre correspondant se trouve dans l'environnement, sinon les matériaux et l'énergie nécessaires à la production de ces enzymes seront gaspillés. Autrement dit, les gènes de ces enzymes de digestion du sucre ne doivent être activés que lorsque ce sucre est présent. Pour assimiler chaque sucre, en règle générale, plusieurs enzymes sont nécessaires, et non une. Leurs gènes sont situés à proximité de la chaîne d'ADN et leur activité est contrôlée par une région régulatrice d'ADN commune au début du premier gène. Un tel groupe de gènes collaboratifs et contrôlés de manière synchrone est appelé «opéron».Schéma de l'opéron lactose L'opéron lactose se compose de trois gènes. Le premier (LacZ) code pour une protéine de transport qui pompe le lactose dans la cellule, et les deux autres (LacI et LacA) codent pour des enzymes, dont le travail conjoint transforme le lactose en «sucre par défaut» - le glucose. Au début de l'opéron lactose, il y a un site de liaison à l'ARN polymérase (promoteur) et un site de liaison aux protéines régulatrices (opérateur).L'activité de l'opéron lactose est contrôlée par deux signaux. Le premier signal, évidemment, est la concentration de lactose. S'il n'y a pas de lactose, les enzymes de son assimilation ne sont pas nécessaires. Le deuxième signal est un peu plus compliqué. Dans une expérience, Jacob et Mono ont cultivé E. coli sur un milieu contenant à la fois du lactose et du glucose. Dans de telles conditions, le bâton consomme d'abord du glucose et son opéron lactose est désactivé. Lorsque le glucose se termine, la croissance des bactéries s'arrête pendant 15 à 20 minutes, puis se poursuit en raison du lactose. Pendant une pause, l'opéron lactose est activé. C'est-à-dire que la deuxième entrée régulatrice désactive l'opéron lactose lorsque la cellule a plus de sucres disponibles que le lactose (l'absorption du glucose ne nécessite aucune enzyme supplémentaire).D'autres expériences ont montré que l'opéron lactose ne répond pas au glucose lui-même. Au lieu de cela, le niveau de faim (plus précisément, la «qualité» des sucres disponibles) à l'intérieur de la cellule d'E. Coli est codé par une substance de signalisation spéciale. Il est appelé «monophosphate d'adénosine cyclique» ou AMPc. Alors que le bâton croît sur le glucose, il n'y a pratiquement pas d'AMPc dans la cellule. S'il n'y a pas de glucose, mais qu'il y a du maltose ou de l'amidon (substances légèrement plus difficiles à assimiler), un peu d'AMPc est produit. S'il n'y a pas de maltose, mais qu'il y a du fructose ou du lactose, le niveau d'AMPc dans la cellule sera plus élevé. S'il n'y a pas de sucres, mais qu'il y a de l'acide lactique - encore plus élevé, et enfin, s'il n'y a rien de savoureux et que vous devez manger de la glycérine, alors le niveau d'AMPc sera le plus élevé. Ainsi, deux signaux contrôlent l'opéron lactose: la concentration de lactose et la concentration d'AMPc.Comment ces signaux affectent-ils l'activité des gènes? Un gène actif est un gène sur lequel l'ARN polymérase produit de nombreux ARN messagers. L'ARN polymérase commence toujours à travailler sur cet opéron en atterrissant sur la même région d'ADN (il est appelé le «promoteur»). Où est la place pour le lactose et l'AMPc? L'ARN polymérase ne peut pas reconnaître la totalité des centaines de signaux qui régulent l'activité de milliers de gènes. Pour cela, il existe des protéines régulatrices spéciales (elles sont appelées «facteurs de transcription»). Deux protéines régulatrices sont impliquées dans la gestion de l'opéron lactose, une pour chaque signal d'entrée: un répresseur lactose et un activateur catabolique.Répresseur de lactose (violet) sur l'ADN Unrépresseur de lactose est une protéine qui peut se lier au lactose ou à une séquence d'ADN spécifique. Il existe deux de ces séquences dans le génome d'Escherichia coli, toutes deux au début de l'opéron lactose, de chaque côté du promoteur. Les molécules de répresseur de lactose adhèrent fermement les unes aux autres en groupes de quatre molécules de protéines. La liaison d'un tel quatre à l'ADN tord le brin d'ADN en une boucle serrée dans laquelle le promoteur finit. L'ARN polymérase ne peut pas s'asseoir sur un tel promoteur tordu et le gène s'éteint. Si le lactose apparaît dans la cellule, il se lie au répresseur du lactose et tombe de l'ADN, libérant le promoteur de l'ARN polymérase - l'opéron commence à fonctionner. C'est ainsi que l'entrée régulatrice de l'opéron en termes de concentration de lactose est arrangée.Un activateur catabolique sur l'ADN Unactivateur catabolique répond de manière similaire à la concentration d'AMPc. Certes, il y a plusieurs différences dans les détails:- l'activateur catabolique se trouve sur l'ADN juste à côté du promoteur et ne le chevauche pas. Au contraire, il améliore la liaison de l'ARN polymérase au promoteur et augmente l'activité de l'opéron.- un activateur catabolique lie l'AMPc et l'ADN. Sans AMPc, il ne peut pas se lier à l'ADN.- un activateur catabolique est nécessaire pour la pleine activité de l'opéron lactose, mais même sans lui, l'opéron fonctionnera encore pour environ 5% de l'activité totale. Le répresseur du lactose régule l'opéron dans une plage de 0 à 100%.- Les molécules de l'activateur catabolique sont connectées par paires, pas par quatre. Dans le prochain article, nous verrons à quoi cette différence conduit.Autrement dit, nous pouvons écrire une table de vérité pour l'opéron lactose:L'opéron lactose fonctionne presque comme un élément logique ET. Seulement 5% de l'activité à l'état "lactose +, AMPc -" sortent légèrement de l'image idéale.Suite avec graphiques et matan: https://geektimes.ru/post/283686/ Source: https://habr.com/ru/post/fr399233/
All Articles