Dans ma publication précédente , deux questions ont été examinées: les principes fondamentaux de la biologie moléculaire et les méthodes de création des constructions dont nous avons besoin pour des gènes basés sur des vecteurs plasmidiques. Maintenant, vous devez comprendre comment introduire un vecteur plasmidique dans une cellule bactérienne, c'est-à-dire pour effectuer une "transformation". Dans le même temps, nous apprendrons quelque chose sur la structure de la membrane bactérienne et comment elle peut être surmontée, ainsi que pourquoi certaines bactéries sont qualifiées de compétentes, comment tout cela est associé aux toxines bactériennes et à la résistance bactérienne aux antibiotiques, et bien d'autres faits intéressants.
Donc, nous avons un vecteur plasmidique sur nos mains et il existe une souche bactérienne adaptée à ce vecteur, il reste à introduire le vecteur dans la bactérie.
Dans un
article précédent , le
travail de Frederick Griffith a été mentionné, ce qui démontre clairement le fait que l'ADN peut pénétrer spontanément dans les cellules bactériennes. La capacité d'une cellule bactérienne à absorber une molécule d'ADN environnementale est appelée «compétence» (ce qui donne lieu à de nombreuses blagues sur le fait que même les cellules unicellulaires in vitro sont compétentes, mais pas votre collègue de laboratoire). Si l'absorption ne se produit pas sous l'influence de certaines conditions créées artificiellement, alors nous parlons de «compétence naturelle». Comment se produit un transport d'ADN similaire à l'intérieur d'une bactérie?
1) La structure de la membrane cellulaire des bactéries
Pour commencer, nous considérons brièvement la structure de la membrane des bactéries gram-positives et gram-négatives.
La coquille d'une bactérie gram-négative se compose de deux bicouches lipidiques: la membrane externe et la membrane interne (l'intérieur est également appelé plasma). Entre eux se trouve l'espace périplasmique rempli de protéines et de peptidoglycanes remplissant des fonctions importantes.
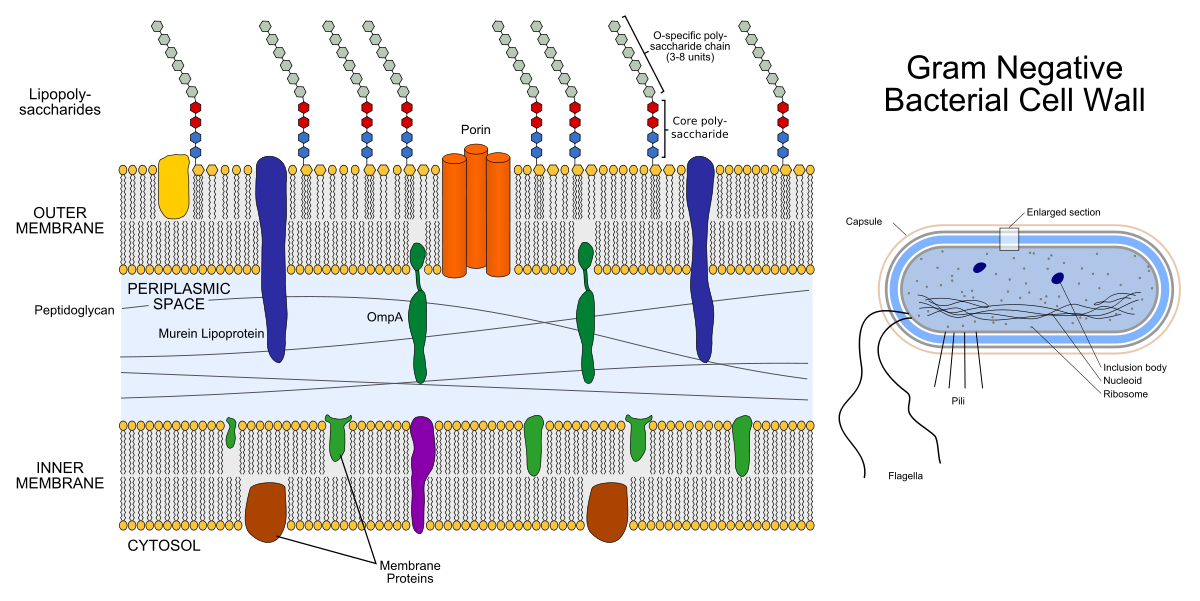 À gauche, la structure de la membrane de la bactérie gram-négative. Les gizmos colorés à l'intérieur des membranes sont des protéines membranaires et les chaînes qui sortent sont des lipopolysaccharides. À droite, la structure de la bactérie gram-négative elle-même.
À gauche, la structure de la membrane de la bactérie gram-négative. Les gizmos colorés à l'intérieur des membranes sont des protéines membranaires et les chaînes qui sortent sont des lipopolysaccharides. À droite, la structure de la bactérie gram-négative elle-même.Le peptidoglycane est un hétéropolymère composé de N-acétylglucosamine réticulée de manière covalente (NAG) et d'acide N-acétylmuramique (NAM). Il s'avère que la chaîne ....- NAG-NAM-NAG-NAM-NAG- .... «Hétéro» dans le mot «hétéropolymère» signifie que le polymère n'est pas constitué des mêmes éléments (comme le polyéthylène), mais de deux éléments différents. L'enzyme de synthèse des peptidoglycanes, la transpeptidase, est une cible pour les antibiotiques β-lactamines (antibiotiques pénicilline et céphalosporines): ils inhibent son activité, du fait du processus de division bactérienne, il n'y a pas assez de peptidoglycane pour les deux cellules filles. La couche de peptidoglycane est également une cible pour l'enzyme lysozyme: elle détruit la liaison covalente entre NAG et NAM. En médecine, le lysozyme est utilisé comme antiseptique, par exemple, il est un composant actif de certains médicaments pour le traitement des maux de gorge, et dans l'industrie alimentaire, il peut être utilisé comme conservateur (complément alimentaire E1105 - c'est le lysozyme).

L'enveloppe des bactéries à Gram positif se compose uniquement d'une membrane interne (plasma), d'un espace périplasmique très mince et d'une couche externe de peptidoglycane plus épaisse que les bactéries à Gram négatif.
Ces obstacles qui entravent tout composé dans le cytoplasme bactérien sont insurmontables pour tout sauf l'eau, ainsi que les petites molécules non polaires et hydrophobes (oxygène et azote moléculaires, dioxyde de carbone et autres). La pénétration de ces composés à travers la coque est appelée "transport passif" ("simple diffusion"), elle se produit toujours le long du gradient de concentration. Les molécules non polaires peuvent passer librement à travers les membranes car le volume interne des membranes est également non polaire (à l'intérieur de la membrane, elles sont constituées de résidus hydrophobes d'acides gras), et les composés non polaires se dissolvent bien dans les solvants non polaires (alcool, acétone, acides gras).
Une image comique dans laquelle l'hydrophobie (hydrophobie) des résidus d'acides gras à l'intérieur d'une membrane à deux couches se joue.À leur tour, les grosses molécules polaires (acides aminés, monosaccharides, nucléotides), les particules chargées (ions) et les macromolécules (ADN, protéines) ne peuvent pas pénétrer à travers la membrane cellulaire comme ça, car elles ne sont bien solubles que dans les solvants polaires (par exemple, dans l'eau) . Alors, comment les molécules d'ADN pénètrent-elles dans une cellule bactérienne?
2) Mécanismes de transport induit de composés dans les cellules depuis l'environnement
Tout d'abord, nous avons entre les mains le fait évident que la bactérie ne peut pas vivre uniquement de l'eau, de l'oxygène et du dioxyde de carbone, pour la vie, elle a besoin de nombreux autres composés. Il existe donc des moyens de délivrer ces composés dans la cellule. Ces méthodes sont la «diffusion facilitée» et le «transport actif».
2.1) Diffusion de la lumière
Un exemple de diffusion facilitée (bien que ce cas particulier, au contraire, soit nocif pour les bactéries) est le transport des ions potassium à travers la membrane d'une cellule bactérienne en combinaison avec la valinomycine, un antibiotique synthétisé par certains champignons. La valinomycine libre ne peut pas pénétrer dans la cellule, car les groupes polaires sont exposés à l'extérieur. L'ion potassium est également un «chemin fermé».
 Une sculpture représentant la structure tridimensionnelle d'un complexe de valinomycine et d'ions potassium (l'ion est représenté comme une boule au centre). Il a été installé devant l'entrée de l'Institut M. M. Shemyakin et Yu. A. Ovchinnikov de chimie bioorganique, Académie russe des sciences (Moscou), dans lequel le mécanisme de l'activité antibactérienne de la valinomycine et la structure tridimensionnelle de son complexe avec l'ion potassium ont été établis.
Une sculpture représentant la structure tridimensionnelle d'un complexe de valinomycine et d'ions potassium (l'ion est représenté comme une boule au centre). Il a été installé devant l'entrée de l'Institut M. M. Shemyakin et Yu. A. Ovchinnikov de chimie bioorganique, Académie russe des sciences (Moscou), dans lequel le mécanisme de l'activité antibactérienne de la valinomycine et la structure tridimensionnelle de son complexe avec l'ion potassium ont été établis.Mais après la formation d'un complexe de valinomycine avec un ion potassium, il change sa structure tridimensionnelle de sorte que toute sa surface extérieure devienne hydrophobe (non polaire), et tous les groupes hydrophiles sont cachés à l'intérieur. Dans ce cas, l'ion potassium est caché à l'intérieur de la molécule de valinomycine; par conséquent, le complexe valinomycine + ion potassium peut pénétrer à travers la membrane. Le complexe peut se décomposer en valinomycine et ion libres à tout moment à l'intérieur et à l'extérieur de la cellule, mais la diffusion généralement facilitée des ions conduit à l'égalisation de leur concentration dans les espaces intracellulaires et extracellulaires, et la cellule contrôle le transport des ions pour une raison: leur concentration élevée fatal. C'est sur cette base que repose l'effet antibactérien de la valinomycine.
Représentation schématique de l'antibiotique fongique valinomycine complexé avec l'ion potassium.Un autre exemple est les pores incontrôlés, qui sont de grandes protéines transmembranaires qui permettent à certaines substances de passer librement à travers la membrane en fonction du gradient. Un exemple d'un tel pore transmembranaire est le
Staphylococcus aureus Staphylococcus aureus α-toxine (un autre nom pour la toxine est α-hémolysine. Il se compose des parties «hémo» et «lysine». «Hemo» signifie globules rouges; «lysine» signifie du mot "lyse", c'est-à-dire "mort cellulaire")
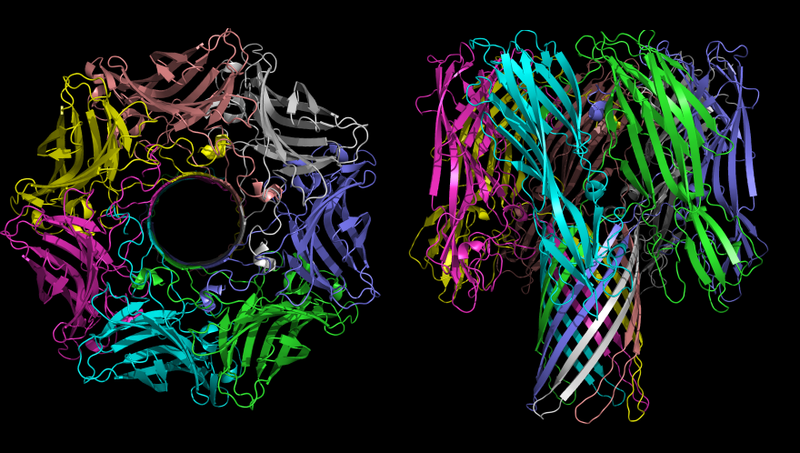 La structure tridimensionnelle de l'oligomère α-hémolysine - la toxine de la bactérie Staphylococcus aureus. Dans l'image de gauche, l'heure est clairement visible.
La structure tridimensionnelle de l'oligomère α-hémolysine - la toxine de la bactérie Staphylococcus aureus. Dans l'image de gauche, l'heure est clairement visible.La fonction principale de l'α-hémolysine dans le processus pathogène est la création de canaux incontrôlés dans la membrane cellulaire, qui, selon les conditions environnementales, peuvent passer des ions monovalents, des ions calcium ou ATP. Comme mentionné ci-dessus, le déséquilibre ionique est extrêmement toxique et le transport de l'ATP
depuis la cellule est nocif, car l'ATP est un lien indispensable dans presque tous les processus chimiques de n'importe quelle cellule.
2.2) Transport actif
Comme on peut le voir dans les exemples ci-dessus, une diffusion facilitée ne nécessite pas d'énergie. Au contraire, le transport actif est l'absorption ou la libération de quelque chose par une cellule avec la dépense d'énergie pour chaque acte de transfert. Cela peut être le transport de grosses molécules ou ions polaires, et le transport d'ions peut être effectué à la fois le long du gradient de concentration et contre lui. Bien sûr, le transport actif est un processus contrôlé dans lequel tout un ensemble de protéines intracellulaires et transmembranaires est impliqué. Ainsi, la cellule doit d'abord dépenser des ressources pour synthétiser toutes ces protéines, puis dépenser à nouveau de l'énergie pour l'utilisation de ce système de transport.
Un exemple de transport actif est le système de transport dépendant de l'énergie des composés hydrophobes et amphiphiles d'une cellule vers l'environnement (efflux).
Le système de transport actif d'un antibiotique d'une cellule bactérienne vers l'extérieur, composé de trois protéines formant un seul complexe transmembranaire. La figure reflète le fait que la protéine de transport d'efflux (violette) peut «attraper» des molécules de médicament directement à la fois dans le cytoplasme et dans la membrane.Ce système présente un intérêt particulier car il provoque la multi-résistance de certaines bactéries aux antibiotiques: la cellule s'en débarrasse avant de pouvoir lui nuire gravement. De plus, plus de la moitié des cas de multirésistance de
Pseudomonas aeruginosa Pseduomonas aeruginosa sont associés à un transport actif d'antibiotiques. Plus important encore, des systèmes d'efflux actifs de «nouveaux» antibiotiques ont déjà été découverts qui sont utilisés contre des bactéries résistantes aux médicaments antibactériens couramment utilisés. Par exemple, le médicament
Linezolid utilisé dans le cas de l'inefficacité des antibiotiques classiques est rejeté par le système d'efflux basé sur les protéines AcrAB et TolC. Un autre fait intéressant est qu'une bactérie intelligente ne gaspille pas d'énergie lors de la synthèse à grande échelle des protéines du système d'efflux dans les situations où elles ne sont pas nécessaires. Mais dès l'apparition de substances nocives pour la cellule, la synthèse est rapidement activée et la bactérie commence à résister activement. Soit dit en passant, l'efflux n'est pas le seul mécanisme de protection des bactéries contre les antibiotiques, j'écrirai également un article sur d'autres méthodes.
La structure du linézolide est un antibiotique utilisé contre les souches résistantes de bactéries gram-positives.3) Compétence des cellules bactériennes
Nous avons donc examiné les mécanismes de transport transmembranaire de composés qui ne peuvent pas pénétrer cette barrière sans assistance extérieure. Dans des conditions naturelles, l'ADN est transporté dans la bactérie uniquement par transport actif en raison de la coopération de tout un spectre de protéines intracellulaires et transmembranaires. La capacité non induite d'une cellule à absorber l'ADN de l'extérieur est appelée «compétence naturelle».
3.1) Compétence naturelle des cellules bactériennes
L'hypothèse la plus étayée de l'émergence de la compétence naturelle des bactéries est l'hypothèse que ce mécanisme permet aux bactéries de survivre dans des conditions extrêmes pour elles (les conditions extrêmes pour une cellule sont appelées «stress»). Le fait est que le stress entraîne souvent des dommages à l'ADN «chromosomique» d'une bactérie, et si une cellule veut survivre, elle doit en quelque sorte «réparer» son «chromosome» (une telle «réparation» est appelée «réparation»). De plus, il est connu que si une séquence d'ADN est introduite dans la cellule qui est similaire à une partie du propre ADN de la cellule, alors il est très probable qu'elle intègre ce «nouveau» morceau dans son génome. Que se passe-t-il dans des conditions stressantes? Certaines cellules meurent et deviennent des donneurs d'ADN pour ceux de leurs frères qui se battent encore pour la vie. En effet, au sein d'une même culture bactérienne, il est très probable que les génomes des représentants soient très similaires, de sorte que les bactéries survivantes peuvent utiliser l'ADN des parents décédés pour réparer leur génome.
De plus, dans des conditions confortables dans la culture cellulaire, seule une fraction d'entre elles a une compétence naturelle, et la proportion de cellules compétentes de leur nombre total est alors la plus élevée, puis la culture est dans une certaine phase de croissance - la phase
logarithmique .
 Une parcelle typique de bactéries en culture au fil du temps. La phase logarithmique est la phase de croissance, suivie d'un plateau et de la phase d'extinction, dans laquelle les cellules manquent déjà de nutriments et les toxines s'accumulent.
Une parcelle typique de bactéries en culture au fil du temps. La phase logarithmique est la phase de croissance, suivie d'un plateau et de la phase d'extinction, dans laquelle les cellules manquent déjà de nutriments et les toxines s'accumulent.Bien entendu, le chercheur souhaite rendre le processus de transfert du vecteur plasmidique dans la cellule aussi efficace que possible: plus le processus de transformation est efficace, moins le vecteur plasmidique est nécessaire pour garantir de bons résultats. Par conséquent, dans des conditions de laboratoire, ils ne dépendent généralement pas de la cellule elle-même, mais créent plutôt des cellules dotées d'une compétence artificielle.
3.2) Compétence artificielle des cellules bactériennes
Il existe deux méthodes principales pour créer la compétence artificielle des bactéries: le traitement d'une culture cellulaire avec des ions divalents suivi d'un chauffage à court terme et la soi-disant électroporation.
3.2.1) Traitement de la culture cellulaire avec des ions divalents, suivi d'un chauffage à court terme.
En règle générale, dans le laboratoire, il y a un réfrigérateur avec une température d'environ -80 degrés Celsius (parfois ils sont appelés "kelvinators" en l'honneur d'une des entreprises manufacturières), bouchés par des tubes à essai avec des cellules en phase
logarithmique . Ceci est fait pour ne pas perdre de temps à les créer chaque fois qu'ils sont nécessaires (et cela prend beaucoup de temps, cela ne fonctionnera pas plus vite qu'en une journée). Et pour que lors du stockage les cellules ne meurent pas à cause du gel, des cryoprotecteurs sont ajoutés au milieu: glycérol, propylène glycol ou diméthylsulfoxyde.
Réfrigérateur de laboratoire selon l'auteur de la photo.Lorsqu'un chercheur doit transformer une souche spécifique, il retire les cellules du Kelvinator et place le tube dans la glace. Ensuite, les cations divalents et le vecteur plasmidique dont nous avons besoin sont ajoutés aux cellules dans la phase
logarithmique de croissance. Le tube à essai ne doit pas quitter la glace. Un donneur typique de tels cations est le chlorure de calcium (CaCl2), dans ce cas, notre cation divalent est Ca2 +. Il n'y a aucune explication sans équivoque de la façon dont le Ca2 + aide l'ADN à pénétrer les bactéries:
- Certaines sources affirment que des concentrations élevées de Ca2 + provoquent des lacunes dans la membrane cellulaire et des dommages à l'ADN, et plus tôt nous avons constaté que les dommages à l'ADN sont un signal pour que la cellule devienne compétente;
- Selon d'autres sources, ces ions servent de «pont» qui attache le vecteur plasmidique à la cellule. Le fait est que les ions chargés positivement se fixent simultanément aux groupes de polysaccharides chargés négativement sur la membrane externe des bactéries et aux groupes chargés négativement d'ADN plasmidique. S'il n'y avait pas d'ions dans le milieu, alors l'ADN serait repoussé de la cellule, et cela est complètement inutile pour le chercheur.
Il est possible que les deux hypothèses soient vraies.
Le mécanisme supposé de participation des ions Ca2 + à la transformation. Premièrement, les ions se fixent aux groupes d'ADN chargés négativement (cercles jaunes) et aux polysaccharides ancrés dans la membrane (cercles jaunes). Ensuite, en raison de l'ion, une sorte de pont se forme: "polysaccharide (-)" - "ion Ca (2+)" - "ADN (-)", qui fixe l'ADN à la surface des cellules.Après une incubation bactérienne à une température proche de zéro, le soi-disant choc thermique est produit - chauffage à court terme de la culture à 42 degrés. En règle générale, la durée du traitement thermique est de 30 à 45 secondes, après quoi le tube à essai contenant des bactéries retourne dans le bain de glace (avec un choc thermique plus long, les cellules peuvent mourir). Les raisons possibles de l'efficacité du choc thermique sont les mêmes mécanismes: une température élevée peut endommager l'ADN et les protéines (ce qui induit la compétence) et peut également créer des «trous» dans la membrane cellulaire à travers lesquels pénètrent des vecteurs plasmidiques attachés à la membrane par les ions calcium.
Ensuite, les cellules sont étalées sur des boîtes de Pétri avec un milieu sélectif et des colonies s'y développent.
3.2.2) Électroporation
La base de la méthode d'
électroporation est le fait que le champ électrique peut endommager la membrane cellulaire. Autrement dit, l'électroporation est la création de pores par un champ électrique.
Tout d'abord, les bactéries et un vecteur plasmidique sont placés dans un tube spécial (cuvette). Ensuite, la cuvette est placée dans un appareil spécial qui, pendant une fraction de seconde, crée une différence de potentiel de 0,1 à 10 kV, ce qui entraîne à son tour des dommages réversibles aux membranes et les vecteurs peuvent pénétrer librement dans les cellules. Après un certain temps, la culture bactérienne est ensemencée sur des boîtes de Pétri avec un milieu sélectif.
Une faible concentration d'ions est une exigence importante pour le milieu liquide dans lequel se trouvent les cellules et le vecteur au moment de l'électroporation: sinon, un court-circuit peut se produire.
Postface
Chaque colonie sur une plaque est la descendance proliférante d'
une seule bactérie (c'est-à-dire que si nous voyons 100 colonies sur une plaque, cela signifie qu'il y avait initialement 100 bactéries viables, qui ont ensuite commencé à se diviser).
Après que les colonies apparaissent sur les boîtes, le chercheur sélectionne l'une des colonies et transfère les cellules de la colonie vers un nouveau milieu nutritif liquide sélectif. Après qu'une nouvelle culture cellulaire "ait grandi", elle est à nouveau plantée sur une nouvelle tasse avec le même milieu nutritif sélectif. En conséquence, nous obtenons une tasse sur laquelle seuls les descendants des bactéries d'une seule colonie se développent (c'est-à-dire qu'ils sont tous les clones d'une seule bactérie). Et seules ces cellules seront utilisées à l'avenir pour obtenir la protéine dont nous avons besoin dans de grands volumes de milieu nutritif sélectif liquide.Eh bien, résumons les résultats intermédiaires:- Nous avons synthétisé le gène dont nous avons besoin;
- Insérez-le dans un vecteur plasmidique approprié;
- Une souche productrice appropriée a été sélectionnée et transformée;
- Vous avez une culture de clone;
- , .
: , . .