
Pendant des milliers d'années, l'humanité a cherché le paradis, voulant devenir comme des oiseaux. De nombreux grands esprits, de Léonard de Vinci aux frères Wright, ont conçu une grande variété d'avions, dont certains sont restés sur papier, tandis que d'autres sont devenus les ancêtres d'avions et d'hélicoptères modernes. Maintenant, voler pour une personne n'est pas difficile - j'ai acheté un billet d'avion et le tour est joué. La plupart d'entre nous ne pensent même pas au fonctionnement de cet énorme oiseau métallique qui peut nous transférer d'un continent à un autre. La vérité entre les avions modernes et les vrais oiseaux n'est pas tellement en commun, mais avec le développement de robots autonomes, la situation est légèrement différente. De nombreux scientifiques tentent de créer un oiseau robot, et pour que cette initiative soit couronnée de succès, il est nécessaire de comprendre non seulement comment les oiseaux volent, mais aussi comment atterrir. Aujourd'hui, nous vous rencontrerons une étude dans laquelle les diplômés de l'Université de Stanford ont étudié en détail le mécanisme de plantation d'un perroquet nommé Gary et ses proches. Comment les oiseaux atterrissent, comment comprennent-ils qu'ils peuvent ou non atterrir sur une surface donnée, et quelle est la cinématique de leurs membres lors de l'atterrissage? Nous trouverons des réponses à ces questions et à d'autres dans le rapport des chercheurs. Allons-y.
Base d'étude
Quelle est la différence entre les avions modernes et les oiseaux en termes d'atterrissage? Le Boeing 777, par exemple, a besoin d'une piste d'une longueur d'au moins 2500 m, un hélicoptère a besoin d'une surface plate (de préférence), comme des hélicoptères. Les oiseaux, quant à eux, ne sont pas limités dans la géométrie de la toile de plantation et peuvent atterrir sur presque toutes les surfaces (si l'on considère les oiseaux dans leur ensemble, plutôt qu'une seule espèce). Si nous parlons de robots autonomes, leurs capacités en termes d'atterrissage sont très limitées. Certains d'entre eux peuvent atterrir avec succès sur des surfaces spécialement conçues, mais cela ne peut être comparé à un pic ordinaire, qui peut atterrir sur des surfaces verticales, inclinées et horizontales, tant qu'il y a des griffes auxquelles s'accrocher.
 C’est pratique pour moi (pic fourmi / Melanerpes formicivorus).
C’est pratique pour moi (pic fourmi / Melanerpes formicivorus).Les chercheurs notent que leurs prédécesseurs ont déjà effectué des études qui ont quantifié la dynamique des pattes et des ailes des oiseaux lors de la plantation. Cependant, on ne sait pas encore comment cette dynamique change en fonction des différents types de surfaces qui servent de site d'atterrissage. Après tout, non seulement la géométrie de la zone d'atterrissage est importante, mais aussi la texture de la surface.
Il est également intéressant de noter que des études antérieures n'ont pas été menées sur des oiseaux vivants; par conséquent, dans la pratique, il n'a pas été possible d'évaluer pleinement la force de préhension des pattes et des griffes. C'est pourquoi certaines questions sont restées éteintes: comment les oiseaux comprennent-ils exactement que le site d'atterrissage convient; quelle est la dynamique des membres au moment de toucher et de saisir; pourquoi pour les oiseaux atterrir sur une variété de surfaces est une question insignifiante, etc.
Dans les travaux que nous envisageons aujourd'hui, les chercheurs ont décidé de tester les mécanismes et la dynamique du débarquement des oiseaux arboricoles en observant les perroquets de Lesson (
Forpus coelestis ).
 Budgie de la leçon.
Budgie de la leçon.Les sujets ont volontairement atterri sur des poteaux de différentes géométries et textures. Au total, il y avait 9 options: 3 «naturelles», 3 avec un diamètre modifié et 3 avec une texture modifiée.
Les options "naturelles" sont nommées ainsi, car les perchoirs étaient faits de véritables rameaux (19 mm de diamètre) de trois arbres différents:
- Chêne de Californie - la surface est rugueuse dans différentes parties d'une branche;
- ceiba est magnifique - la surface est relativement lisse, les branches sont molles;
- osmanthus parfumé - la surface est rugueuse longitudinalement, les branches sont dures.
Pour les expériences avec un diamètre modifié du poteau, des broches de bouleau ordinaires de différents diamètres ont été utilisées comme matériau:
- 38 mm - un diamètre trop grand pour ce type de perroquet;
- 19 mm - le perroquet peut effectuer une prise partielle (les doigts avant et arrière ne se touchent pas);
- 6 mm - le perroquet peut effectuer une prise complète (les doigts avant et arrière se touchent).
Pour les expériences avec différentes textures, un poteau en bouleau d'un diamètre de 19 mm a été enveloppé dans trois matériaux différents:
- mousse - texture douce et élastique;
- téflon - texture glissante;
- Le papier de verre est une texture rugueuse et rugueuse.
Le perroquet teste les 9 options pour le site d'atterrissage.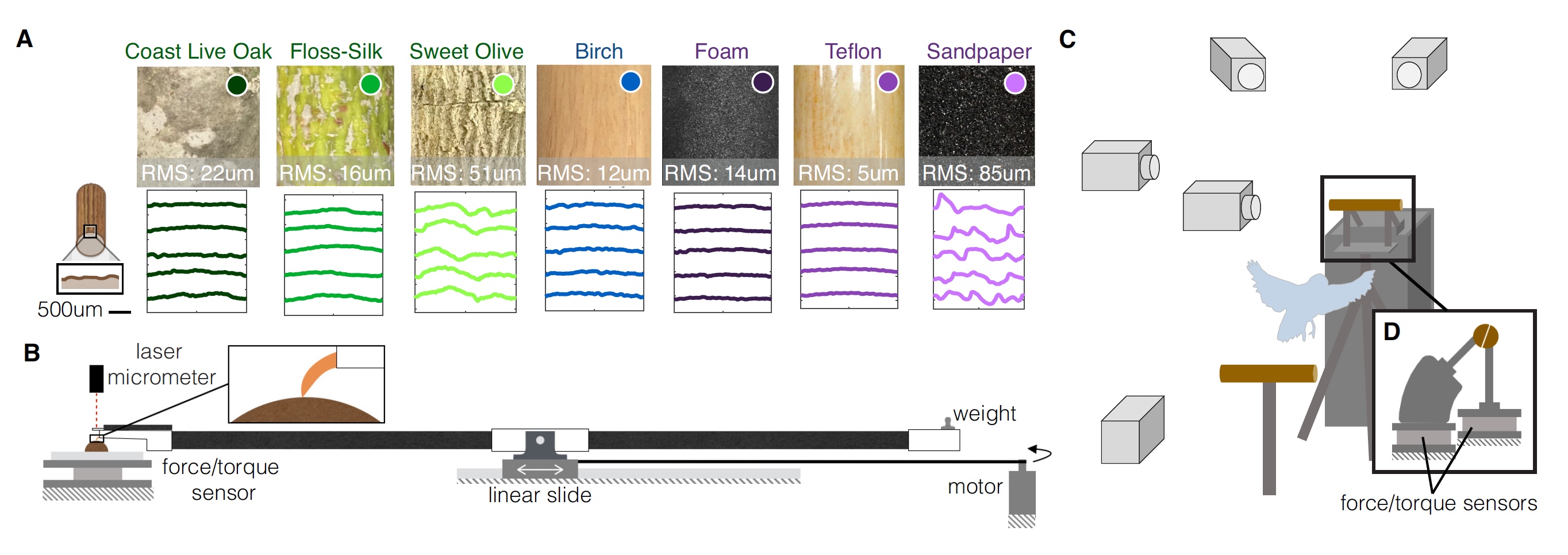 Image n ° 1
Image n ° 1Un balayage tridimensionnel en lumière structurale de chacune des variantes de perforation a été réalisé, ce qui a permis de décrire quantitativement la texture de surface de chacune d'elles et de recréer des profils de surface (
1A ). Pour évaluer l'effet de ces caractéristiques texturales sur l'interaction de la surface de la perche et du pied du perroquet, les chercheurs ont effectué des tests de résistance des doigts et des griffes pour mesurer les forces de friction pour chaque surface. La pression des griffes a également été évaluée pour mesurer la déformation de la surface (
1B ).
Sur le diagramme
1C, on voit que tous les tests ont été effectués sous le regard de plusieurs caméras rapides, et les pôles ont été sciés en deux. Des capteurs de force et de moment de force sont attachés à chaque moitié.
Résultats de recherche
Tout d'abord, considérez la dynamique des ailes et des pattes lors de l'atterrissage.
Dans tous les tests avec toutes les variantes de la surface d'atterrissage, les sujets ont montré la même dynamique classique. L'atterrissage commence par le freinage des ailes (phase d'air), après quoi les pattes absorbent l'impulsion restante au contact du perchoir (phase d'absorption). Le contact a été établi avec les deux pattes avec un délai entre elles de quelques millisecondes, soit le premier pied en contact avec le poteau est le pied préféré:
- Sujet n ° 1 - dans 100% des cas, la patte droite;
- Sujet n ° 2 - dans 83% des cas, le pied gauche;
- Sujet n ° 3 - dans 85% des cas, le pied gauche.
Après le contact, la phase de fixation commence, lorsque le perroquet fixe ses doigts autour du perchoir de manière plus fiable et plus ferme. Cela est suivi d'une étape qui n'est pas toujours trouvée, mais souvent, c'est la phase d'ajustement, quand ils changent en quelque sorte la position des pattes ou des doigts sur la perche.
Toutes les phases d'atterrissage d'un perroquet (correspond au schéma sur 2A ). Image n ° 2
Image n ° 2Parfois, les perroquets, surestimant ou sous-estimant apparemment les conditions du test, ne survolaient / ne survolaient pas légèrement le site d'atterrissage (
2B ), ce qui conduisait à une plus grande dispersion de l'angle d'inclinaison du pied, auquel ils établissent une adhérence statique.
Il est curieux que s'il y a une certaine variation dans les indicateurs de la force des jambes (
2B ) et de la direction (
2C ), les valeurs moyennes de la force d'atterrissage restent étonnamment les mêmes pour différents pôles.
La différence d'indicateurs constatée lors de l'atterrissage s'explique par la stratégie d'atterrissage des perroquets. Il existe une théorie selon laquelle les oiseaux contrôlent leur atterrissage en évaluant visuellement le temps de contact avec la surface,
τ (
t ). Pour un atterrissage réussi, ils ajustent leur vitesse d'approche pour maintenir une constante
τ (
t ). Si un peu plus détaillé:
τ est défini comme la distance au perchoir (
s ) divisée par la vitesse d'approximation (
v ).
Si l'oiseau ralentit avec une décélération constante (
a ), alors
τ =
s /
v = 0,5
à 2 /
at = 0,5
t , auquel cas
τ (
t ) = 0,5.
Si
τ (
t ) <0,5, l'inhibition diminuera jusqu'au moment du contact avec le perchoir. Et si 0,5 <
τ (
t ) <1, le freinage augmentera jusqu'à ce que l'oiseau effectue une collision contrôlée avec la surface d'atterrissage.
Compte tenu de la cinématique décrite ci-dessus, les perroquets maintiennent des valeurs relativement constantes de
τ (
2E ) correspondant aux valeurs des collisions contrôlées (
2F ). La plus petite valeur moyenne
τ = 0,80 a été observée dans les tests avec un poteau avec le plus petit diamètre (6 mm). Et la valeur la plus élevée de
τ = 0,94 était dans les tests avec un poteau recouvert de mousse. Cela suggère que les perroquets augmentent le freinage à l'approche d'une surface d'atterrissage douce.
La présence d'un
τ élevé (c'est-à-dire supérieur à 0,5) dans tous les tests avec tous les types de surface suggère que les oiseaux n'ont pas terminé l'atterrissage au moment du toucher.
Ayant un peu familiarisé avec la dynamique de l'atterrissage, nous allons passer à la cinématique des pieds et des griffes de nos amis à plumes.
Compte tenu de l'uniformité du comportement lors de l'atterrissage, quel que soit le type de surface, les chercheurs ont conclu que les oiseaux réagissent aux différences de texture et de géométrie du pôle d'atterrissage avec précision avec leurs pattes et leurs griffes, et non avec leurs ailes. Cela semble logique, non? Mais, surprise, ce n'est pas entièrement vrai.
La cinématique du pied a montré un ensemble stéréotypé (identique pour tous les types de perchoirs) d'étapes d'atterrissage (
3A ,
3B ).
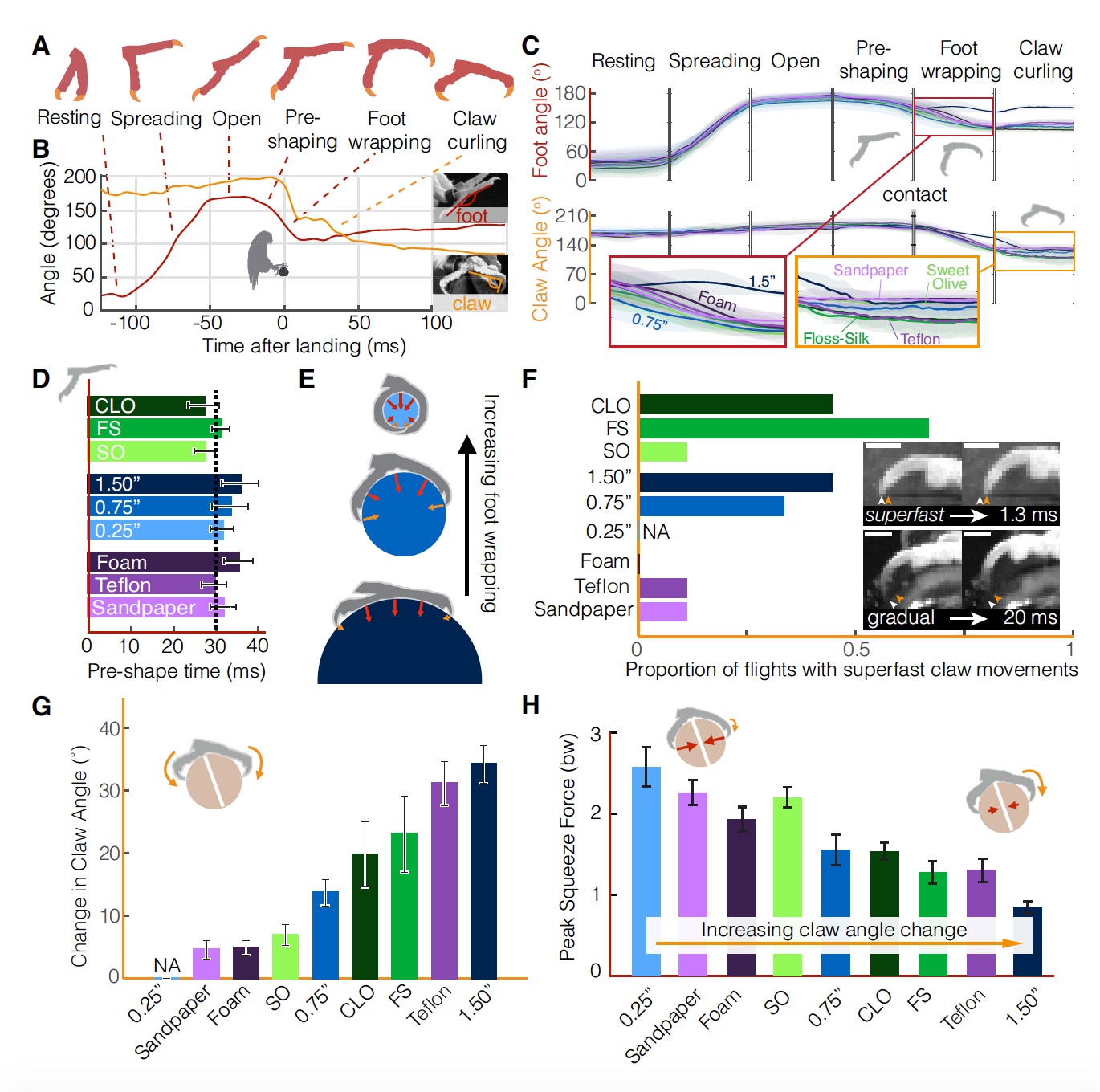 Image n ° 3
Image n ° 3Pendant le vol, les pieds sont dans un état fermé (phase de repos), puis ils commencent à s'ouvrir environ 100 ms avant de toucher le perchoir (phase d'ouverture d'une durée de 40 ± 8 ms), puis il y a une phase "ouverte", quand dans les 21 ± 7 ms l'arrêt entièrement divulgués.
Immédiatement avant le contact, les doigts commencent à se rapprocher (phase de la formation préalable de la capture, durée 31 ± 10 ms). Après avoir établi le contact avec la surface, les doigts commencent à saisir le perchoir (phase de circonférence, durée 19 ± 7 ms). La phase finale est la capture des griffes d'une durée de 185 ± 11 ms.
Les différences de texture et de géométrie du pôle affectent exclusivement la cinématique des paliers après contact avec la surface (
3C ). Jusqu'au contact, la cinématique du pied et de la griffe reste inchangée dans tous les tests, même lorsque les pieds commencent à se fermer pendant la phase de pré-adhérence.
En d'autres termes, si les oiseaux réagissaient à la différence de géométrie et de texture de la surface d'atterrissage, des changements cinématiques seraient observés même pendant les phases avant le contact, c'est-à-dire toujours dans l'air. Au lieu de cela, les perroquets commencent séquentiellement à pré-capturer 30 ms avant l'atterrissage (
3D ). Cela conduit à l'hypothèse qu'ils ont des solutions prévisibles au problème de l'atterrissage sur une surface inhabituelle.
Il est important pour l'oiseau de maintenir un grand angle entre les griffes jusqu'au moment du contact afin d'éviter leur endommagement. De plus, le degré de capture est déterminé par le diamètre du pôle (
3E ). La capture complète se produit dans le cas de pôles d'un diamètre de 6 mm, c'est-à-dire les perroquets peuvent utiliser pleinement toute la force de leurs doigts et de leurs griffes pour se stabiliser.
Il est important de noter qu'au moment du contact, les griffes et / ou les pattes peuvent glisser hors des pôles, en raison de la surface lisse. Par conséquent, l'angle de la griffe peut varier. Étant donné que l'étape de saisie des griffes peut durer plus de 100 ms, les mouvements individuels d'une seule griffe (
3F ) peuvent se produire incroyablement rapidement (1-2 ms).
Les mouvements de griffes ultra-rapides ne sont probablement pas régulés uniquement par la contraction musculaire, suggèrent les chercheurs. Il est également surprenant que parmi les vertébrés, les mouvements musculaires les plus rapides soient 5 à 10 fois plus lents que ces mouvements de griffes ultra rapides (par exemple, les muscles du bras humain - 50 ms, le muscle pectoral du colibri - 8 ms). De cela, nous pouvons conclure que la vitesse de déplacement de la griffe d'un perroquet dépend de la libération d'énergie stockée dans le tendon élastique et de la faible inertie de la griffe. Ces mouvements rapides des griffes permettent aux perroquets d'adapter instantanément leur adhérence en fonction de la géométrie et de la texture de la surface d'atterrissage.
Quant au degré de capture des griffes, il dépend du diamètre du pôle et de la texture (
3G ). L'adhérence la plus prononcée est observée dans le cas d'un diamètre de 38 mm. Il convient de préciser qu'il s'agit simplement de capturer les griffes, et pas toutes les jambes entièrement. Autrement dit, plus le diamètre est grand, plus vous devez vous fier aux griffes, car les pattes n'aideront pas (
3C ,
3E ). L'angle de la griffe par rapport à la surface dépend fortement de sa texture. Par exemple, dans le cas de surfaces plus lisses (Téflon), les griffes se «plient» davantage (
3H ) que dans le cas d'une surface rugueuse (papier de verre), car dans ce cas, elles peuvent simplement compresser la surface un peu plus, assurant ainsi une adhérence fiable. De façon exagérée, les griffes se plient plus lorsque la surface est plus difficile à comprimer.
 Image n ° 4
Image n ° 4Ensuite, les chercheurs ont décidé de vérifier comment, du point de vue de la mécanique, les indicateurs de la résistance à la compression des doigts et de la flexion des griffes changent, et comment ils affectent l'adaptation de l'oiseau à l'une ou l'autre surface lors de l'atterrissage.
Des tests avec différentes textures des pôles ont montré que le bout des doigts (
4A ) et les griffes (
4C ) présentent une
force de cisaillement similaire
* aux moments de glissement et de préhension.
Force de cisaillement * - forces inégales poussant une partie du corps dans une direction particulière et la seconde partie du corps dans le sens opposé.
Pour les patins, la méthode la plus appropriée pour représenter la force de cisaillement en fonction de la force normale pour toutes les surfaces est le modèle de friction Coulomb (
4B ), qui permet de prédire la direction et l'ampleur de la force de friction entre deux corps en contact avec des surfaces sèches (ce modèle est donc également appelé «modèle de friction sèche» )
Pour les griffes (
4C ), le coefficient de frottement peut être 8 fois supérieur à celui des patins (
4A ). Une telle différence peut être causée, entre autres, par l'orientation de la griffe par rapport à la surface (
4D ): sur la surface la plus molle, la force de friction reste relativement constante, mais sur une surface rugueuse, cette force augmente également avec l'augmentation de l'angle de la griffe. Cependant, même aux mêmes angles, les mesures montrent que le coefficient de force maximum pour les griffes est toujours plus élevé que pour les patins (
4E ).
Pour compenser la nature stochastique de l'adhésion de l'embrayage, les oiseaux peuvent profiter de l'équilibrage de la charge et du mouvement des griffes à la surface. La répartition de la charge entre plusieurs griffes réduit efficacement la distribution de la résistance totale attendue de toutes les griffes (
4E ). De plus, les oiseaux peuvent allonger leurs griffes, ce qui leur permet d'augmenter la probabilité d'adhésion à une surface inégale, le cas échéant. Cela augmente la valeur attendue de la force de frottement (
4F ), ce qui explique la flexion plus prononcée des griffes sur des surfaces plus difficiles à comprimer fermement (
3F ). L'étirement des griffes donne un avantage dans le cas de surfaces sur lesquelles les irrégularités sont petites, mais elles sont assez grandes. Cependant, cette affirmation est légèrement incompatible avec les observations, car les perroquets n'ont pas beaucoup cherché à étendre leurs griffes, ce qui suggère qu'ils ne choisissent pas les meilleures bosses lors de la plantation. Par conséquent, ils plient leurs griffes autant que possible jusqu'à ce qu'une force suffisante soit atteinte pour maintenir une prise stable. Si au cours de ce processus, cette force n'est pas atteinte, l'oiseau commencera à ajuster la position des griffes ou à décoller.
 Image n ° 5
Image n ° 5La différence entre les coefficients de frottement des griffes et des patins peut également s'expliquer par la géométrie de ces parties du corps et la géométrie de la surface d'atterrissage avec laquelle elles sont en contact. Les patins ont une plus grande surface et une texture douce, ce qui vous permet de répartir la charge à de nombreux points de contact, ce qui conduit à une force de friction plus uniforme. Les griffes, à leur tour, sont grossières et ont une petite surface, elles dépendent donc d'une interaction directe avec la géométrie de la surface, c'est-à-dire sur l'engrenage des bosses.
Pour quantifier les effets géométriques, les scientifiques ont caractérisé la forme de la griffe dans le plan sagittal en largeur en fonction de la hauteur de la pointe de la griffe (
5A ). En plus d'utiliser les irrégularités de surface existantes, les griffes peuvent également générer des forces de friction à partir de la déformation de surface.
Pour simuler cette interaction, des mesures de la profondeur de pénétration de la griffe dans diverses surfaces (
5B ) et des mesures de la géométrie de la griffe elle-même ont été combinées. La géométrie de la pointe de la griffe a été modélisée par une sphère d'un rayon de 50 μm pour des charges allant jusqu'à 25% du poids corporel (
5C ). Ainsi, il a été constaté que, par exemple, la profondeur de pénétration dans le perchoir du chêne de Californie est de 10 μm, soit 0,2% de la longueur de l'arc externe de la griffe (environ 5 mm).
Afin d'évaluer plus en détail l'effet de la taille des griffes sur la traction, un autre modèle a été créé avec des griffes de différents diamètres de pointe s'accrochant à différentes surfaces (
5D ). Comme prévu, en l'absence de toute pénétration de griffe dans la surface, les surfaces plus rugueuses sont plus appropriées pour une capture réussie (
5E ).
Lorsque des effets de pénétration de surface (
5F ) ont été ajoutés au modèle, il a été constaté que la surface utile était augmentée. De plus, plus l'effet est prononcé, plus l'angle formé entre l'axe vertical et le vecteur de force nette sur la griffe est petit. Si le rayon et la masse de la pointe de la griffe sont mis à l'échelle isométrique et que les forces appliquées dépendent de la masse, le coefficient de profondeur (profondeur de pénétration divisée par le rayon de la pointe de la griffe) sera également mis à l'échelle isométrique.
De cela, nous pouvons conclure logiquement que de petites irrégularités sur cette surface sont suffisantes pour que les petits animaux adhèrent efficacement à la surface.
 Image n ° 6
Image n ° 6En combinant tous les modèles ci-dessus pour divers effets, forces et autres choses, les chercheurs ont créé un modèle bidimensionnel unique, qui prend simultanément en compte les paramètres des griffes, des pattes et du corps dans son ensemble (
6A ).
L'ensemble des données sur la mécanique de contact des pieds et de la surface d'atterrissage a permis d'appliquer une optimisation limitée pour déterminer «l'espace de capture» tridimensionnel de l'oiseau et la combinaison nécessaire de force et de moment de force (
6B ). , , : , . , (
6C ).
, . , . , . : , .
, , . , , , . , .
.
Épilogue
, , . . , . , , , . , . - -, - — . , , - , . , , , .
Vendredi hors-dessus:
( , ), .
Merci de votre attention, restez curieux et passez un bon week-end à tous, les gars! :)
, . ? ? Soutenez-nous en passant une commande ou en le recommandant à vos amis, une
réduction de 30% pour les utilisateurs Habr sur un analogue unique de serveurs d'entrée de gamme que nous avons inventés pour vous: Toute la vérité sur VPS (KVM) E5-2650 v4 (6 cœurs) 10 Go DDR4 240 Go SSD 1 Gbps à partir de 20 $ ou comment diviser le serveur? ( RAID1 RAID10, 24 40GB DDR4).
Dell R730xd 2 ? 2 Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 $199 ! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — $99! . c Dell R730xd 5-2650 v4 9000 ?