
La relation entre partenaires, remplie de soins, signes d'attention et d'empathie, les poètes appellent l'amour, mais les biologistes - une relation inter-sexuelle visant à la survie et à la procréation. Certaines espèces préfèrent en prendre en quantité - se reproduire avec autant de partenaires que possible afin d'augmenter la progéniture, augmentant ainsi les chances de survie de l'espèce entière. D'autres créent des couples monogames qui ne peuvent cesser d'exister qu'après la mort de l'un des partenaires. Pendant de nombreuses années, les scientifiques ont estimé que la première option était beaucoup plus rentable, mais ce n'est pas entièrement vrai. Les couples monogames, en règle générale, élèvent conjointement une progéniture, c'est-à-dire le protéger des prédateurs, obtenir de la nourriture et acquérir certaines compétences, tandis que dans les relations polygames, tout cela repose le plus souvent sur les épaules fragiles des femelles. Bien sûr, il y a des exceptions, mais aujourd'hui ce n'est pas leur sujet. Pendant longtemps, les biologistes se sont intéressés à un autre moment intéressant: les mâles continuent de montrer des signes d'attention aux femelles, même lorsque leur couple est déjà formé et existe depuis plusieurs années. Quelle est la raison de ce comportement, quels en sont les avantages et quels sont les aspects évolutifs associés à cela? Nous trouverons des réponses à ces questions dans le rapport du groupe de recherche. Allons-y.
Base d'étude
Compte tenu du sujet de recherche, nous ne nous concentrerons pas sur les espèces d'oiseaux polygames, mais sur les romantiques à plumes qui tombent amoureux une fois pour toutes.
En parlant de monogamie, il convient de noter qu'il en existe plusieurs types, selon la durée: une saison, plusieurs années et à vie.
Chez les oiseaux, la monogamie saisonnière est la plus courante. Un exemple frappant est celui des oies sauvages. Les femelles nichent et font éclore des œufs, et le mâle garde le territoire. Le deuxième jour après l'éclosion, la famille se rend au plan d'eau le plus proche, où les oisons apprennent à se nourrir. En cas de danger de dépassement sur l'eau, la femelle protège violemment la progéniture, mais le mâle, se souvenant apparemment de questions importantes, fuit le plus souvent. Pas la relation la plus idéale, de toute façon.
 Famille d'oies sauvages.
Famille d'oies sauvages.Si nous parlons de relations, dont la base est la constance, les cigognes sont les meilleures en la matière. Ils créent un couple monogame à vie et ne changent même pas de lieu de résidence sans besoin particulier. Un nid de cigognes, qui peut peser jusqu'à 250 kg et atteindre 1,5 m de diamètre, les dessert pendant de nombreuses années, si les catastrophes naturelles ou l'intervention humaine ne le détruisent pas. En République tchèque, un nid a été créé en 1864.
 Les compétences de construction des cigognes n'ont pas besoin d'être évaluées lorsque vous voyez de telles structures.
Les compétences de construction des cigognes n'ont pas besoin d'être évaluées lorsque vous voyez de telles structures.Contrairement aux oies sauvages, les cigognes ont des responsabilités égales: les deux partenaires éclosent, cherchent de la nourriture, apprennent à leur progéniture à voler et le protègent des dangers. Un rôle important dans les relations des cigognes est joué par différents types de rituels: chant, danse, etc. Le plus curieux est que ces rituels sont effectués non seulement lors de la formation du couple (à la première date), mais tout au long de la vie ensemble (même en remplaçant la femelle pendant l'incubation, le mâle exécute une petite danse). Pour nous, cela a l'air très mignon, romantique et absolument illogique, car d'un point de vue biologique, un tel comportement n'a aucun avantage. C'est bien ça? Et ici, vous pouvez facilement commencer à considérer l'étude elle-même, qui était censée répondre à cette question.
Les éthologues * croient que la manifestation constante par les mâles de leurs sentiments est associée à la préservation de l'état reproducteur chez les femelles.
Ethologie * - une science qui étudie les comportements génétiquement déterminés, c'est-à-dire instincts.
Dans le même temps, on ne sait pas pourquoi un tel comportement dure non seulement pendant la période d'accouplement initiale, mais toute la vie, car il serait plus logique que les mâles investissent plus de force et d'énergie dans la progéniture, plutôt que de manifester des sentiments pour la femelle. À ce jour, de nombreux chercheurs pensaient que l'intensité de l'expression de l'affection pour la femelle affectait directement la qualité de l'accouplement et, par conséquent, la progéniture (c'est-à-dire le nombre d'œufs pondus).
 Un mâle de l'une des espèces d'oiseaux de paradis danse devant une femelle. Comme nous le voyons, le mâle est beaucoup plus brillant que la femelle.
Un mâle de l'une des espèces d'oiseaux de paradis danse devant une femelle. Comme nous le voyons, le mâle est beaucoup plus brillant que la femelle.Cette théorie est confirmée par des observations. La femelle, dont la partenaire est un bel homme non écrit et le premier dépliant du village, met plus d'efforts dans la progéniture que si le mâle n'est ni poisson ni viande. Cela semble drôle et drôle, mais les rituels que les mâles exécutent devant les femelles visent à démontrer non seulement la beauté, mais aussi la force. Il se trouve que le plumage brillant, le beau chant et d'autres manifestations d'attention de la part des mâles ne sont pour les femelles que des signaux cognitifs qu'il décode en informations sur le mâle.
Les scientifiques des universités de Caroline du Nord et de Chicago, dont nous examinons les travaux aujourd'hui, pensent que ce comportement des mâles vise à optimiser le comportement des femelles par rapport au processus de reproduction.
Le modèle proposé par les scientifiques est basé sur de nombreuses expériences qui ont montré que l'amplification de ces signaux par les mâles augmente la contribution des femelles au processus de procréation. Il est supposé que la source de ces effets stimulants sont des réponses perceptives résultant des propriétés de l'environnement, des signaux et du système nerveux en tant que tels. Actuellement, environ 100 exemples de telles «déviations» des systèmes sensoriels conventionnels (audition, vision et odorat) sont connus.
Lorsque le mâle démontre à nouveau ses avantages par rapport aux autres mâles, cela peut affecter positivement le mâle lui-même (la femelle le choisira à coup sûr). Mais pour la femelle, cela peut être un inconvénient, car cela réduira la future mise en œuvre de la reproduction. En d'autres termes, nous avons une situation de «dépassement des attentes». Le mâle, qui est bien meilleur que les autres mâles et montre constamment des signes d'intérêt pour la femelle, recevra l'accouplement et la procréation souhaités, ou plutôt une sorte. Une femme, qui s'attend à un comportement similaire de la part d'autres hommes, mais ne le reçoit pas, peut se retrouver dans une situation déplorable. Un cas similaire est appelé par les scientifiques comme un conflit inter-sexuel: la démonstration par les hommes de beaux augmente dans la population, et chez les femmes la résistance à cette tactique augmente.
Ce conflit a été modélisé à l'aide d'une approche informatique (réseaux de neurones). Dans les modèles obtenus, le dispositif de signalisation (source de signal - mâle) utilise la perception perceptuelle du récepteur (récepteur de signal - femelle), qui stimule les signaux eux-mêmes au détriment de la perception. À un certain moment, un changement dans la perception des signaux dans la population féminine se produit (une sorte de mutation), à partir de laquelle la force du signal de la source (masculine) diminuera considérablement. L'augmentation progressive de ces changements conduira au fait qu'un type particulier de signal sera complètement inefficace. Au fur et à mesure que de tels changements se produisent, certains signaux disparaissent, perdent de leur force, mais de nouveaux apparaissent et le processus recommence.
Ce système très tordu est assez simple en pratique. Imaginez qu'un mâle apparaisse avec une plume brillante (une seule), il se démarque des autres, et les femelles lui donnent la préférence. Puis un mâle apparaît avec deux plumes brillantes, puis avec trois, etc. Mais la force d'un tel signal, compte tenu de sa croissance et de sa propagation, commence à diminuer proportionnellement. Et puis tout à coup un mâle apparaît qui sait chanter magnifiquement et construire des nids. En conséquence, le beau plumage en tant que signal cesse d'être efficace et commence à dégénérer.
Cependant, il y a toujours une exception à la règle - certains conflits inter-sexuels peuvent se transformer en une coopération inter-sexuelle à part entière et très efficace.
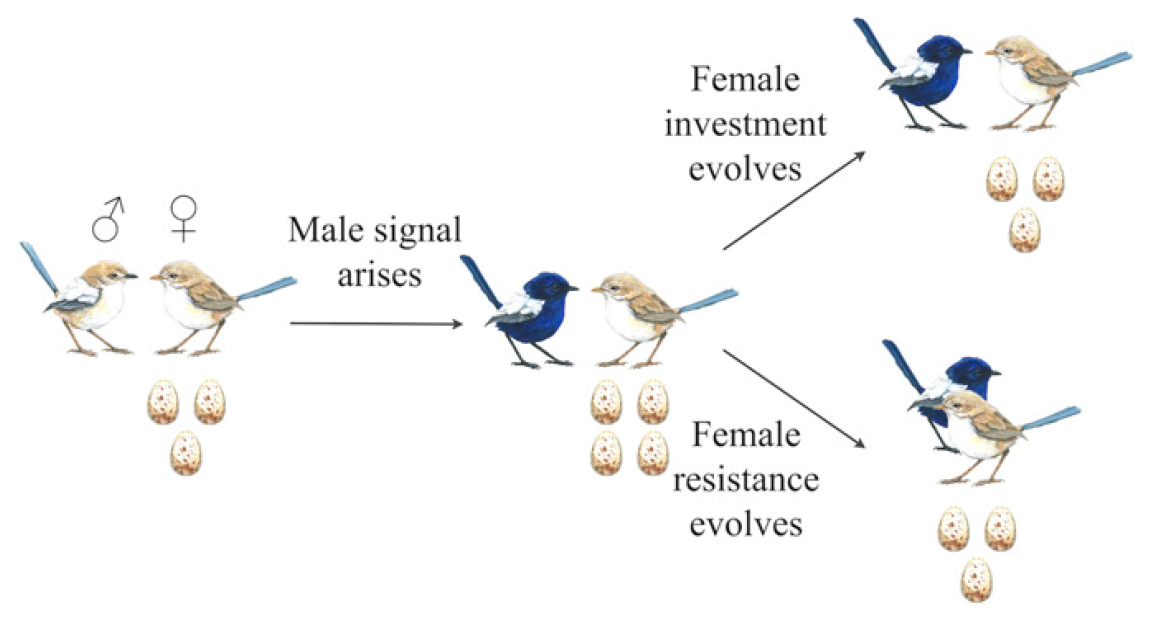 Schéma de l'occurrence d'un conflit inter-sexuel et d'une coopération inter-sexuelle.
Schéma de l'occurrence d'un conflit inter-sexuel et d'une coopération inter-sexuelle.L'essentiel est que le mâle avec un signal plus prononcé oblige la femelle à pondre non pas trois œufs, mais quatre. C'est bon pour le mâle - il aura plus de progéniture avec son patrimoine génétique. Pour une femelle - pas vraiment, car elle devra consacrer plus d'énergie à faire en sorte que tous les petits survivent et atteignent un âge indépendant. Par conséquent, les femelles commencent à se développer parallèlement aux mâles afin d'être plus résistantes à leurs signaux. Le résultat peut être de deux manières: conflit ou coopération.
Dans le cas de la coopération, les femelles évoluent de manière à pondre 3 œufs, comme avant l'apparition d'un signal plus fort des mâles, mais continuent de répondre à ces signaux. Ici, vous avez des astuces pour femmes dans le monde naturel. Ainsi, non seulement une paire est formée, mais une paire qui se soutient mutuellement au niveau optimal pour la procréation en termes d'interaction signal-réponse.
Les hommes ne peuvent pas évoluer en arrière, en gros. Leurs signaux amplifiés vers les femelles donnent le résultat sous la forme d'une maçonnerie de trois œufs, c'est-à-dire pas comme prévu. Cependant, la réduction du signal au niveau précédent sera également inefficace, car elle entraînera une diminution du nombre d'oeufs dans l'embrayage à deux. Il en résulte un cercle vicieux - les mâles ne peuvent pas réduire la force du signal et ne peuvent pas l'augmenter, car les femelles dans le premier cas donneront moins de progéniture, et dans le second elles ne répondront pas.
Naturellement, ni les hommes ni les femmes n'ont l'intention ou le désir malveillant de s'asservir. L'ensemble de ce processus se déroule au niveau génétique et vise uniquement le bénéfice de la progéniture d'une seule paire et le bien-être de l'espèce dans son ensemble.
Résultats de recherche
En utilisant la modélisation mathématique, les scientifiques ont évalué les conditions dans lesquelles une coopération entre les sexes peut se produire. Une caractéristique quantitative avec une valeur moyenne de
z f décrit la principale contribution de la femelle à sa progéniture. Initialement, la valeur moyenne peut évoluer vers sa valeur optimale
z opt , qui dépend de deux variables: le bénéfice de la contribution (le nombre de descendants survivants) et le coût de la contribution pour les femelles (
c f ). La dernière variable est estimée après la reproduction, ce qui implique que certaines femelles survivent et peuvent à nouveau produire une progéniture l'année prochaine, ce qui entraîne une augmentation du nombre de générations.
Dans cette étude, plusieurs termes seront souvent utilisés qui méritent une petite clarification:
- signaux - la manifestation de l'attention de la part des hommes à l'adresse des partenaires féminines (chant, danse et autres rituels) se déroulant en paires formées;
- contribution / investissement - la réponse des femelles à ces signaux, qui se manifeste par un plus grand nombre d'oeufs dans l'embrayage, plus de temps pour s'occuper de la future progéniture, etc.
- répondant - femme répondant aux signaux masculins;
- coûts - le coût de la contribution des femelles à la progéniture (temps dans le nid, temps pour chercher de la nourriture, état de santé en raison du plus grand / plus petit nombre d'œufs dans la ponte, etc.).
De nouveaux signaux masculins et des réponses féminines à ces signaux ont été modélisés à l'aide de modificateurs à locus unique diallyle à recombinaison libre, combinant ainsi des approches génétiques quantitatives et génétiques de population. Au
locus * , qui contrôle la réponse de la femelle (A), une fréquence élevée de l'allèle
répondeur * (A2) est initialement observée, ce qui correspond à la perception perceptuelle existante
Locus * - localisation d'un gène spécifique sur la carte génétique du chromosome.
Les allèles * sont différentes formes du même gène situées aux mêmes loci de chromosomes homologues. Les allèles déterminent la voie de développement d'un trait particulier.
Le gène répondeur * (Rsp) est un gène associé fonctionnellement à un facteur de perturbation de ségrégation (gène SD), dont l'allèle actif (Rsp +) est capable de supprimer l'expression de SD.
Le locus de signal (B) est initialement fixé pour l'allèle sans signal (B1). Ensuite, l'allèle B2 est introduit, ce qui provoque l'apparition de signaux provenant des mâles.
La manifestation des signaux pour les hommes a également un prix (
s m ), mais augmente la contribution de la partenaire féminine (A2) de α. Par exemple, α peut être exprimé comme un œuf supplémentaire dans l'embrayage. Dans le même temps, une augmentation de la contribution de la femelle peut également se manifester sous la forme des effets positifs qu'elle exerce sur sa progéniture.
Par conséquent, la paire dans laquelle le mâle porte l'allèle de signalisation et la femelle porte l'allèle répondeur (c'est-à-dire les paires A2B2) a une contribution supplémentaire du côté féminin et, par conséquent, est plus fertile que les 3 autres combinaisons.
 Variantes de combinaisons de mâles et de femelles par le rapport des signaux et des réponses à ceux-ci.
Variantes de combinaisons de mâles et de femelles par le rapport des signaux et des réponses à ceux-ci.Le nombre de descendants qui ont survécu pour se reproduire l'année suivante est affecté
par la dépendance à la densité * à l'intérieur de la couvée et la dépendance à la densité de la couvée après le plumage.
Dépendant de la densité * - Les processus dépendants de la densité se produisent lorsque les taux de croissance de la population sont contrôlés par la densité de cette population.
Un autre groupe de variables est associé à la mortalité des femelles et des mâles après la naissance de la progéniture. Ces variables sont déterminées par la contribution à la couvée (
c m est la contribution des mâles,
c f est la contribution des femelles), les coûts du signal pour les mâles (
s m ) et la mortalité non sélective (
d m sont des mâles et
d f sont des femelles).
Les veuves, les veufs, les mineurs et toute personne auparavant célibataire se réunissent pour former de nouveaux couples, et le cycle annuel se termine. Dans le modèle étudié, l'accent est mis sur la monogamie génétique, par conséquent, tous les types de sélection sexuelle (c'est-à-dire la concurrence entre les individus pour un partenaire) sont exclus des calculs.
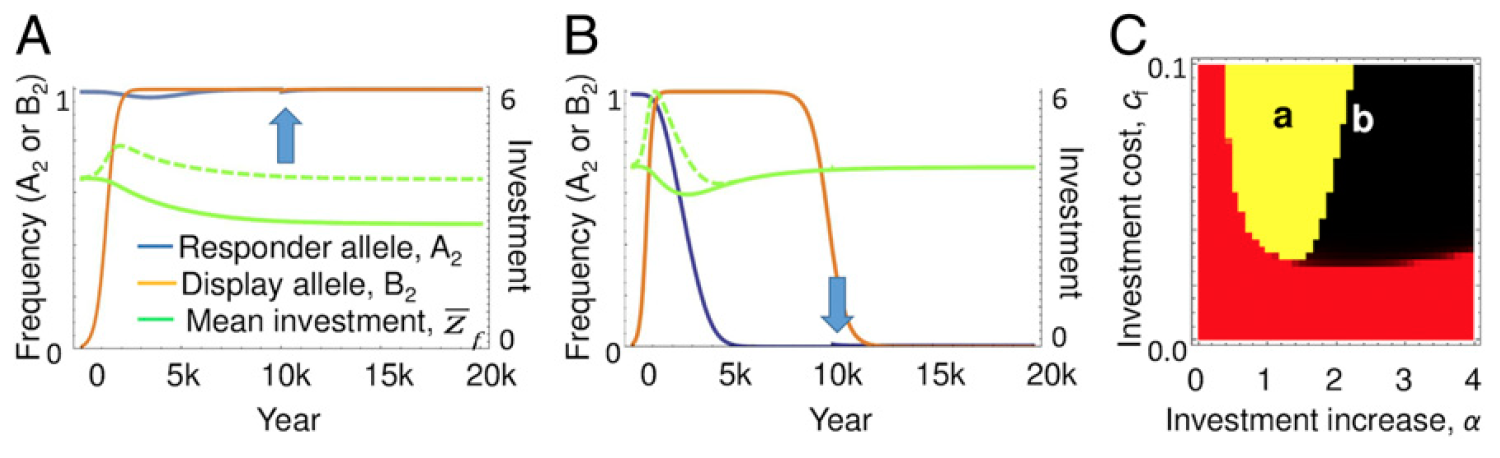 La relation entre l'évolution des signaux, les répondants et les entrées.
La relation entre l'évolution des signaux, les répondants et les entrées.La modélisation a montré qu'un équilibre stable est atteint lorsque les mâles donnent des signaux et que les femelles y répondent. À l'équilibre, la contribution entière à la progéniture est rétablie au niveau qui était avant l'apparition de signaux supplémentaires de mâles.
La figure
A ci-dessus montre un exemple de dynamique évolutive, où la contribution des femelles à la progéniture revient au niveau optimal, qui est le résultat de l'évolution du caractère quantitatif de la contribution (la ligne verte en pointillés est la contribution réelle, et la ligne verte continue est la contribution qui n'a pas été réalisée en raison du manque de réponse femelles pour des signaux supplémentaires de mâles). Le graphique
B montre un autre exemple où un conflit entre les sexes entraîne la perte du répondant.
Et sur le graphique
C, deux paramètres sont sélectionnés qui affectent ce résultat: une augmentation de la contribution causée par des signaux supplémentaires (
α ), et le coût des femelles pour cette contribution (
c f ). Dans la zone rouge du graphique, les signaux n'augmentent jamais, car leur valeur dépassera le profit. Dans les zones jaunes et noires, la fréquence du signal augmente, ce qui entraîne une augmentation des investissements coûteux de la part des femmes. Dans la région jaune, la réponse est en réduisant le caractère d'investissement quantitatif, ce qui conduit à une fixation constante des allèles des signaux et des répondants. Dans la région noire, où les femelles réagissantes ont plus d'investissements induits, l'allèle correspondant est rapidement perdu, suivi de signaux, comme dans les modèles traditionnels de conflit entre les sexes (figure
B ).
La frontière verticale entre les régions rouge et jaune est le point auquel les mâles reçoivent une contribution supplémentaire à la progéniture du fait que les femelles équilibrent les coûts de leur production de signal. La bordure horizontale séparant les régions jaune et noire du rouge apparaît de manière similaire, mais pour une raison moins évidente. Lorsque les coûts des femelles pour la contribution (
c f ) sont faibles, la valeur optimale de la contribution (
z opt ) sera relativement élevée et, par conséquent, la contribution des femelles sera beaucoup plus importante dans les conditions initiales. La conséquence en est que les signaux donnent au mâle un avantage proportionnellement plus faible de la contribution qu'il provoque, qui est encore compensée par ses coûts.
L'espace des paramètres dans lequel les signaux et les réponses sont fixés (jaune) varie en fonction de la force de sélection et de la dispersion génétique de l'allèle répondant. Par exemple, lorsque la fréquence initiale de l'allèle répondant est de 0,9 au lieu de 0,99, comme le montre l'image n ° 2, l'introduction de signaux conduit à une sélection plus efficace pour les répondeurs (la variance génétique initiale est plus élevée) et la région noire s'étend à gauche.
Des signaux masculins peuvent apparaître même s'ils impliquent des coûts qui réduisent la contribution du mâle à la couvée actuelle (paramétrée par
s fec ), affectant ainsi directement la
forme physique * du mâle et de la femelle, plutôt que de réduire la probabilité de survie du mâle.
Fitness * - la capacité de reproduire des individus avec un certain génotype.
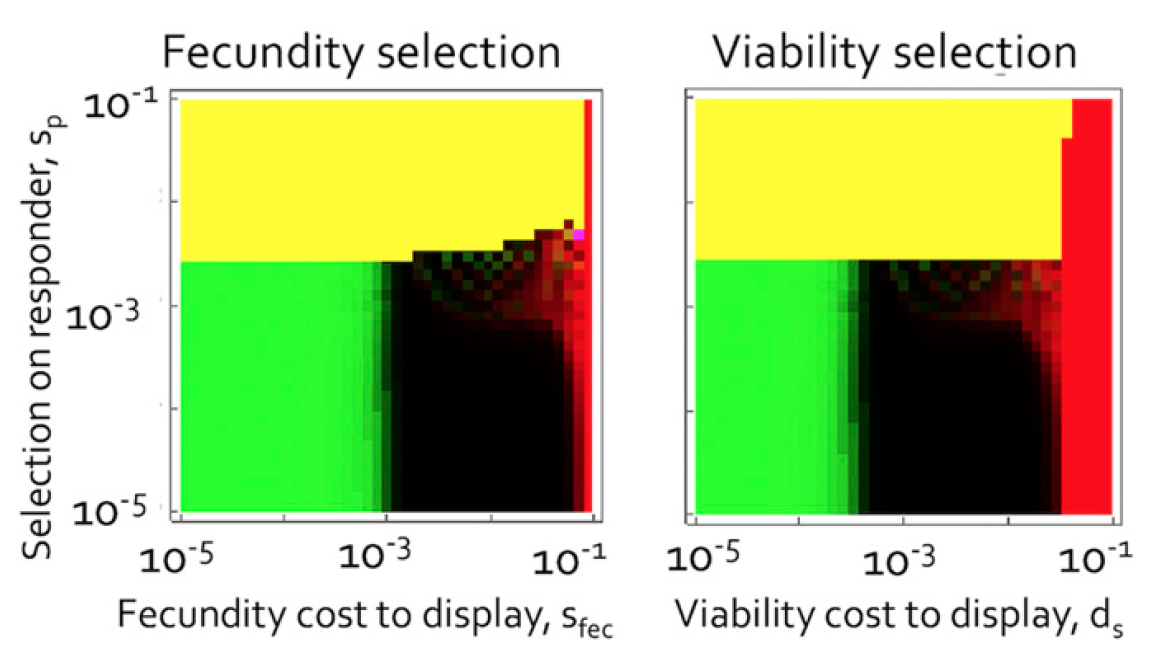 Le rapport des coûts et des signaux de fertilité (à gauche) et le rapport des coûts et des signaux de vitalité.
Le rapport des coûts et des signaux de fertilité (à gauche) et le rapport des coûts et des signaux de vitalité.Dans l'aspect de la fertilité, lorsque les signaux mâles sont fixes (zone jaune), tous les mâles mettent moins dans la progéniture qu'avant l'apparition des signaux. Dans ce cas, la contribution des femelles sera plus importante qu'elle ne l'était avant la manifestation des signaux des mâles.
La plus grande contribution des femelles, lorsque les coûts des mâles sont régulés par la fécondité (plutôt que par la viabilité), augmente le nombre moyen de descendants par couple, mais cela ne compense pas entièrement. Au fil du temps, la plus grande contribution des femelles entraîne une augmentation du nombre moyen de descendants qui ont atteint le plumage, mais diminue la viabilité moyenne des femelles. Cela conduit à la formation d'un nouvel équilibre entre ces deux forces, où le nombre moyen de descendants est inférieur à celui de la viabilité normale ou aux conditions initiales (avant la manifestation des signaux).
: 1% ( ), 1.3%, 0.5%, 0.16%.
(, - ), , .. . , .
(, , ..). , , , , , .
Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de consulter le
rapport des scientifiques et
les documents supplémentaires qui s'y rapportent.
. , : , . , , . , . , , . , , , . . , . 3 , , .
, — . , .
, , , , .
, . , , , .
- 2.0:
() , (BBC Earth, — ).
Merci de votre attention, restez curieux et passez un bon week-end à tous, les gars! :)
Merci de rester avec nous. Aimez-vous nos articles? Vous voulez voir des matériaux plus intéressants?
Soutenez-nous en passant une commande ou en le recommandant à vos amis, une réduction de 30% pour les utilisateurs Habr sur un serveur d'entrée de gamme analogique unique que nous avons inventé pour vous: Toute la vérité sur VPS (KVM) E5-2650 v4 (6 cœurs) 10 Go DDR4 240 Go SSD 1 Gbps à partir de 20 $ ou comment diviser le serveur? (les options sont disponibles avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).
Dell R730xd 2 fois moins cher? Nous avons seulement
2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV à partir de 199 $ aux Pays-Bas! Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - à partir de 99 $! Pour en savoir plus sur la
création d'un bâtiment d'infrastructure. classe utilisant des serveurs Dell R730xd E5-2650 v4 coûtant 9 000 euros pour un sou?