 Dalam pekerjaan pada simulator sistem saraf, sejauh ini kita hanya berurusan dengan aspek-aspek pekerjaan yang dipelajari dengan baik. Tetapi kompleksitas pemodelan sistem saraf dan alasan mengapa kecerdasan buatan belum diciptakan adalah kurangnya pemahaman yang lengkap tentang cara kerja sel saraf. Banyak proses yang terjadi dalam sel saraf dan sistem saraf secara keseluruhan dijelaskan secara rinci, tetapi tidak ada algoritma yang jelas untuk pekerjaan mereka yang dapat ditransfer ke model atau program komputer.Gagasan sederhana dari algoritma neuron membantu memecahkan masalah ini.Isi1. Simulator sistem saraf. Bagian 1. Penambah sederhana2. Simulator sistem saraf. Bagian 2. Modulasi neuroelement3. Simulator sistem saraf. Bagian 3. Neuroelement asosiatif4. Memori, konsolidasi memori, dan neuron nenek.5. Pemodelan emosi atau rasa baru secara elektronik6. Otak kecil yang menakjubkan7. Struktur dan pengaturan otak mulaiSaya suka analogi menyusun teori kerja otak dengan mengumpulkan teka-teki dari buku Jeff Hawkins, On Intelligence. Dalam menyusun puzzle ini, kita tidak memiliki beberapa elemen dan beberapa elemen dari puzzle lain, tetapi kita memiliki sejumlah besar data pada sistem saraf dan otak, yang berarti kita memiliki puzzle yang hampir berkumpul, sehingga kita dapat secara kasar membayangkan keseluruhan gambar, dan menggunakan imajinasi kita mengidentifikasi unsur-unsur yang hilang.Tujuan saya adalah untuk membuat model logis dari sistem saraf, kita dapat mengatakan pembuatan sketsa dari apa yang digambarkan pada teka-teki yang belum selesai, apalagi, itu harus sesuai dan tidak bertentangan dengan semua elemen puzzle yang tersedia dan pada saat yang sama secara logis lengkap. Untuk mengisi kekosongan, beberapa landasan teori diciptakan, yang mungkin tampak kontroversial bagi sebagian orang. Tetapi untuk model pada tahap ini, hal utama adalah memungkinkan Anda untuk meniru fenomena yang diamati baik internal maupun eksternal yang terjadi di sistem saraf. Dalam kerangka model yang diperoleh, banyak fenomena dapat dijelaskan, seperti memori dan konsolidasi memori, emosi, spesialisasi neuron, dan banyak lagi.Pada bagian kedua, kami menemukan bahwa ada tiga jenis kegiatan refleks yang didirikan oleh akademisi I.P. Pavlov. Jika semuanya sangat jelas dengan mekanisme biologis kecanduan dan kepekaan, maka pembentukan refleks yang dikondisikan tidak sesederhana kelihatannya. Faktanya adalah bahwa manifestasi eksternal dari mekanisme ini dipelajari dan dijelaskan secara luas, tidak ada penjelasan tentang bagaimana ini terjadi pada tingkat sel.
Dalam pekerjaan pada simulator sistem saraf, sejauh ini kita hanya berurusan dengan aspek-aspek pekerjaan yang dipelajari dengan baik. Tetapi kompleksitas pemodelan sistem saraf dan alasan mengapa kecerdasan buatan belum diciptakan adalah kurangnya pemahaman yang lengkap tentang cara kerja sel saraf. Banyak proses yang terjadi dalam sel saraf dan sistem saraf secara keseluruhan dijelaskan secara rinci, tetapi tidak ada algoritma yang jelas untuk pekerjaan mereka yang dapat ditransfer ke model atau program komputer.Gagasan sederhana dari algoritma neuron membantu memecahkan masalah ini.Isi1. Simulator sistem saraf. Bagian 1. Penambah sederhana2. Simulator sistem saraf. Bagian 2. Modulasi neuroelement3. Simulator sistem saraf. Bagian 3. Neuroelement asosiatif4. Memori, konsolidasi memori, dan neuron nenek.5. Pemodelan emosi atau rasa baru secara elektronik6. Otak kecil yang menakjubkan7. Struktur dan pengaturan otak mulaiSaya suka analogi menyusun teori kerja otak dengan mengumpulkan teka-teki dari buku Jeff Hawkins, On Intelligence. Dalam menyusun puzzle ini, kita tidak memiliki beberapa elemen dan beberapa elemen dari puzzle lain, tetapi kita memiliki sejumlah besar data pada sistem saraf dan otak, yang berarti kita memiliki puzzle yang hampir berkumpul, sehingga kita dapat secara kasar membayangkan keseluruhan gambar, dan menggunakan imajinasi kita mengidentifikasi unsur-unsur yang hilang.Tujuan saya adalah untuk membuat model logis dari sistem saraf, kita dapat mengatakan pembuatan sketsa dari apa yang digambarkan pada teka-teki yang belum selesai, apalagi, itu harus sesuai dan tidak bertentangan dengan semua elemen puzzle yang tersedia dan pada saat yang sama secara logis lengkap. Untuk mengisi kekosongan, beberapa landasan teori diciptakan, yang mungkin tampak kontroversial bagi sebagian orang. Tetapi untuk model pada tahap ini, hal utama adalah memungkinkan Anda untuk meniru fenomena yang diamati baik internal maupun eksternal yang terjadi di sistem saraf. Dalam kerangka model yang diperoleh, banyak fenomena dapat dijelaskan, seperti memori dan konsolidasi memori, emosi, spesialisasi neuron, dan banyak lagi.Pada bagian kedua, kami menemukan bahwa ada tiga jenis kegiatan refleks yang didirikan oleh akademisi I.P. Pavlov. Jika semuanya sangat jelas dengan mekanisme biologis kecanduan dan kepekaan, maka pembentukan refleks yang dikondisikan tidak sesederhana kelihatannya. Faktanya adalah bahwa manifestasi eksternal dari mekanisme ini dipelajari dan dijelaskan secara luas, tidak ada penjelasan tentang bagaimana ini terjadi pada tingkat sel.
 Sebagai contoh, kita tahu bahwa ketika aktivitas dua pusat saraf digabungkan dengan waktu, busur refleks terbentuk di antara mereka. Yaitu selanjutnya, setelah aktivasi satu pusat saraf, akan ada transfer eksitasi ke pusat saraf lain. Jika kita membagi busur refleks secara kiasan menjadi segmen, dan menganggap segmen tersebut sebagai elemen individual. Dapat dikatakan bahwa ketika busur refleks dari refleks terkondisi terbentuk, perpindahan arah terjadi di setiap segmen. Setiap segmen memilih arah tertentu di mana ia akan, transmisi eksitasi saraf terjadi ketika diaktifkan. Tentu saja, perlu dicatat bahwa arah ini tidak didefinisikan dengan jelas untuk segmen, tetapi dapat dikorelasikan dalam nilai-nilai tertentu.Anda bahkan dapat berbicara tentang penguatan transmisi ke arah tertentu dan melemahnya ke arah lain.
Sebagai contoh, kita tahu bahwa ketika aktivitas dua pusat saraf digabungkan dengan waktu, busur refleks terbentuk di antara mereka. Yaitu selanjutnya, setelah aktivasi satu pusat saraf, akan ada transfer eksitasi ke pusat saraf lain. Jika kita membagi busur refleks secara kiasan menjadi segmen, dan menganggap segmen tersebut sebagai elemen individual. Dapat dikatakan bahwa ketika busur refleks dari refleks terkondisi terbentuk, perpindahan arah terjadi di setiap segmen. Setiap segmen memilih arah tertentu di mana ia akan, transmisi eksitasi saraf terjadi ketika diaktifkan. Tentu saja, perlu dicatat bahwa arah ini tidak didefinisikan dengan jelas untuk segmen, tetapi dapat dikorelasikan dalam nilai-nilai tertentu.Anda bahkan dapat berbicara tentang penguatan transmisi ke arah tertentu dan melemahnya ke arah lain. Ketika refleks diperkuat oleh pengulangan berulang, kita dapat berbicara tentang memperbaiki dan memperkuat transmisi dalam arah untuk setiap segmen. Konsep ini mengarah pada kesimpulan bahwa jika kita membagi seluruh korteks menjadi segmen yang sama, kita akan mengamati orientasi tertentu di setiap arah dengan akurasi dan kekuatan yang berbeda. Setiap segmen akan dikeringkan oleh bagian dari busur refleks dari refleks terkondisi atau tidak terkondisi. Agaknya, orientasi dalam proses pembelajaran ini dapat disempurnakan atau diubah.
Ketika refleks diperkuat oleh pengulangan berulang, kita dapat berbicara tentang memperbaiki dan memperkuat transmisi dalam arah untuk setiap segmen. Konsep ini mengarah pada kesimpulan bahwa jika kita membagi seluruh korteks menjadi segmen yang sama, kita akan mengamati orientasi tertentu di setiap arah dengan akurasi dan kekuatan yang berbeda. Setiap segmen akan dikeringkan oleh bagian dari busur refleks dari refleks terkondisi atau tidak terkondisi. Agaknya, orientasi dalam proses pembelajaran ini dapat disempurnakan atau diubah. Jika kita beralih ke paradigma saraf, maka itu tidak memberikan orientasi ke arah. Kami memiliki membran dan dendrit yang menerima sinyal dan akson, di mana sinyal ditransmisikan lebih lanjut ke sel lain setelah penjumlahan ruang-waktu, yaitu, sinyal ditransmisikan dalam satu arah sepanjang akson ke ujungnya. Tetapi pada saat yang sama, kita masih mengamati pembentukan distribusi eksitasi terarah di otak, dengan pembentukan refleks terkondisi.
Jika kita beralih ke paradigma saraf, maka itu tidak memberikan orientasi ke arah. Kami memiliki membran dan dendrit yang menerima sinyal dan akson, di mana sinyal ditransmisikan lebih lanjut ke sel lain setelah penjumlahan ruang-waktu, yaitu, sinyal ditransmisikan dalam satu arah sepanjang akson ke ujungnya. Tetapi pada saat yang sama, kita masih mengamati pembentukan distribusi eksitasi terarah di otak, dengan pembentukan refleks terkondisi.Paradigma neuron, . . -, , .. . -, - -. -, . , — , . , . , , . .
Jika kita menganggap bukan sel tunggal sebagai unit fungsional dari directional switching, tetapi area kecil sel, maka kita dapat melihat bahwa sel dan prosesnya saling terkait erat, dan dalam arah yang berbeda. Ini memberikan elemen komunikasi terarah dengan banyak input dan output di berbagai arah. Bentuk neuron disebabkan oleh perubahan evolusioner. Bentuk sel dibentuk dalam sistem saraf di mana hanya fungsi aktivitas saraf yang paling sederhana dilakukan. Ketika perkembangan kehidupan di Bumi membutuhkan penambahan pembentukan refleks tangkapan ke rangkaian fungsi sistem saraf, maka evolusi tidak mengikuti jalur restrukturisasi sel, tetapi peningkatan jumlah mereka dan padatnya proses jalinan proses mereka.Dengan demikian, properti directional switching didistribusikan dalam kelompok neuron, dalam perubahan kekuatan sinapsisnya. Sebuah neuroelement asosiatif adalah unit fungsional dalam pemodelan dan oleh karena itu analog dalam biologi untuk itu adalah sekelompok neuron yang fenomena pengalihan arah akan diekspresikan.Kami menemukan bahwa arah propagasi eksitasi penting bagi kami, tetapi bagaimana penentuan arah ini untuk setiap elemen fungsional. Diketahui bahwa eksitasi cenderung menyebar ke sumber eksitasi lainnya, dan fokus eksitasi yang lebih kuat dan lebih luas menarik yang lebih lemah (kesimpulan I. Pavlov). Yaitu jika elemen fungsional menerima kegembiraan, maka entah bagaimana ia harus menentukan arah yang selanjutnya akan dibentuk dan dipertahankan dalam strukturnya.Dalam pekerjaan saya pada pemodelan, saya mulai dari ide interaksi elektromagnetik sel-sel saraf , dan gagasan ini memberikan jawaban untuk banyak teka-teki tentang otak, memberi teori dan model yang menjelaskan banyak aspek sistem saraf.
Bentuk neuron disebabkan oleh perubahan evolusioner. Bentuk sel dibentuk dalam sistem saraf di mana hanya fungsi aktivitas saraf yang paling sederhana dilakukan. Ketika perkembangan kehidupan di Bumi membutuhkan penambahan pembentukan refleks tangkapan ke rangkaian fungsi sistem saraf, maka evolusi tidak mengikuti jalur restrukturisasi sel, tetapi peningkatan jumlah mereka dan padatnya proses jalinan proses mereka.Dengan demikian, properti directional switching didistribusikan dalam kelompok neuron, dalam perubahan kekuatan sinapsisnya. Sebuah neuroelement asosiatif adalah unit fungsional dalam pemodelan dan oleh karena itu analog dalam biologi untuk itu adalah sekelompok neuron yang fenomena pengalihan arah akan diekspresikan.Kami menemukan bahwa arah propagasi eksitasi penting bagi kami, tetapi bagaimana penentuan arah ini untuk setiap elemen fungsional. Diketahui bahwa eksitasi cenderung menyebar ke sumber eksitasi lainnya, dan fokus eksitasi yang lebih kuat dan lebih luas menarik yang lebih lemah (kesimpulan I. Pavlov). Yaitu jika elemen fungsional menerima kegembiraan, maka entah bagaimana ia harus menentukan arah yang selanjutnya akan dibentuk dan dipertahankan dalam strukturnya.Dalam pekerjaan saya pada pemodelan, saya mulai dari ide interaksi elektromagnetik sel-sel saraf , dan gagasan ini memberikan jawaban untuk banyak teka-teki tentang otak, memberi teori dan model yang menjelaskan banyak aspek sistem saraf.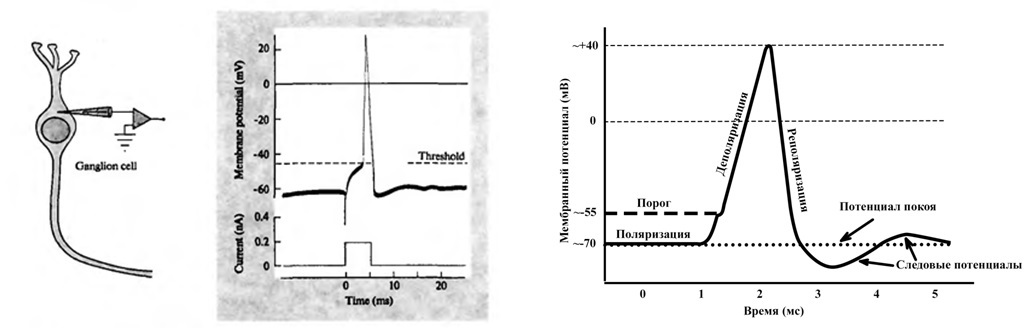 Impuls saraf di seluruh sistem saraf memiliki bentuk yang sama, dan dengan analogi dengannya, neuroelement asosiatif memiliki sifat muatan yang menandai perubahan dalam muatan total pada permukaan membran unit fungsional. Yaitu hukum perubahan tertentu dari karakteristik tertentu yang disebut tuduhan diberikan.
Impuls saraf di seluruh sistem saraf memiliki bentuk yang sama, dan dengan analogi dengannya, neuroelement asosiatif memiliki sifat muatan yang menandai perubahan dalam muatan total pada permukaan membran unit fungsional. Yaitu hukum perubahan tertentu dari karakteristik tertentu yang disebut tuduhan diberikan.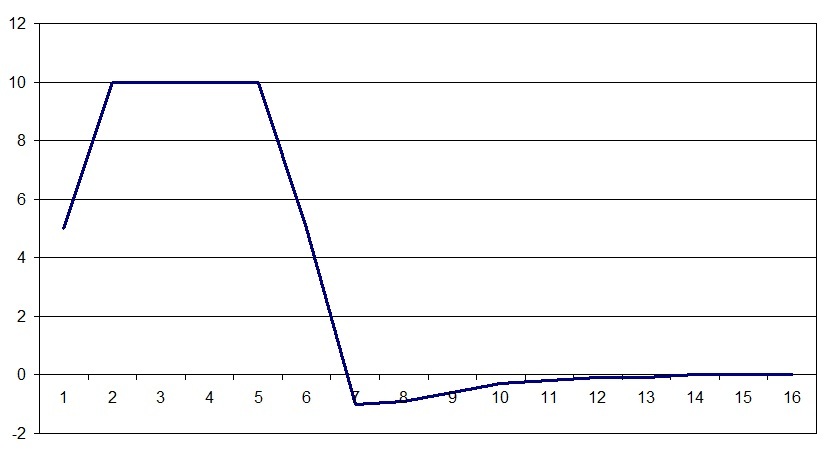 Jadi hukum diatur dalam program, skala horizontal adalah waktu dalam seperseratus detik, muatan vertikal dalam satuan relatif. Ini agak berbeda dari jadwal lonjakan di bagian maksimum lebih lama. Ini disebabkan oleh fakta bahwa nilai-nilai lonjakan ditentukan pada satu titik di jaringan saraf selama perjalanan eksitasi, dan grafik muatan adalah cerminan muatan di atas semua permukaan sel atau kelompok sel. Juga, keadaan sisa jaringan saraf diambil sebagai nol pada skala muatan. Perlu dicatat bahwa hukum perubahan muatan juga mencerminkan potensi jejak, yang sebelumnya dianggap sebagai konsekuensi dari osilasi tertentu atau penyelarasan muatan yang dipisahkan oleh membran, tetapi perilaku muatan ini sangat penting bagi model.
Jadi hukum diatur dalam program, skala horizontal adalah waktu dalam seperseratus detik, muatan vertikal dalam satuan relatif. Ini agak berbeda dari jadwal lonjakan di bagian maksimum lebih lama. Ini disebabkan oleh fakta bahwa nilai-nilai lonjakan ditentukan pada satu titik di jaringan saraf selama perjalanan eksitasi, dan grafik muatan adalah cerminan muatan di atas semua permukaan sel atau kelompok sel. Juga, keadaan sisa jaringan saraf diambil sebagai nol pada skala muatan. Perlu dicatat bahwa hukum perubahan muatan juga mencerminkan potensi jejak, yang sebelumnya dianggap sebagai konsekuensi dari osilasi tertentu atau penyelarasan muatan yang dipisahkan oleh membran, tetapi perilaku muatan ini sangat penting bagi model. Gambar di atas menunjukkan diagram neuroelement asosiatif. Sinyal dari sinapsis kerja langsung (X1, X2, X3 ... Xn) masuk ke adder (a). Dan jika jumlah yang dihasilkan melebihi batas tertentu (b), maka neuroelement akan diaktifkan. Ketika neuroelement diaktifkan, biayanya akan mulai berubah sesuai dengan hukum yang berlaku (c). Informasi tentang perubahan ini dan lokasi elemen itu sendiri akan tersedia untuk seluruh sistem. Selanjutnya, pada titik waktu tertentu, mekanisme untuk menentukan vektor dari arah propagasi yang disukai (g) dimulai. Ini terjadi dengan memperoleh posisi muatan rata-rata tertentu dari semua neuroelements aktif, yaitu pusat massa muatan, ditandai dengan titik di ruang angkasa. Titik ini disebut titik pola,karena untuk setiap kombinasi sel aktif dan status muatannya pada saat yang dihitung dalam waktu untuk setiap neuroelement, posisi titik ini akan berbeda. Sederhananya, muatan neuroelements mempengaruhi penentuan vektor arah dari propagasi eksitasi yang disukai, muatan positif menarik eksitasi, muatan negatif menolak.Untuk menentukan vektor propagasi eksitasi yang disukai, aturan dipilih: di
Gambar di atas menunjukkan diagram neuroelement asosiatif. Sinyal dari sinapsis kerja langsung (X1, X2, X3 ... Xn) masuk ke adder (a). Dan jika jumlah yang dihasilkan melebihi batas tertentu (b), maka neuroelement akan diaktifkan. Ketika neuroelement diaktifkan, biayanya akan mulai berubah sesuai dengan hukum yang berlaku (c). Informasi tentang perubahan ini dan lokasi elemen itu sendiri akan tersedia untuk seluruh sistem. Selanjutnya, pada titik waktu tertentu, mekanisme untuk menentukan vektor dari arah propagasi yang disukai (g) dimulai. Ini terjadi dengan memperoleh posisi muatan rata-rata tertentu dari semua neuroelements aktif, yaitu pusat massa muatan, ditandai dengan titik di ruang angkasa. Titik ini disebut titik pola,karena untuk setiap kombinasi sel aktif dan status muatannya pada saat yang dihitung dalam waktu untuk setiap neuroelement, posisi titik ini akan berbeda. Sederhananya, muatan neuroelements mempengaruhi penentuan vektor arah dari propagasi eksitasi yang disukai, muatan positif menarik eksitasi, muatan negatif menolak.Untuk menentukan vektor propagasi eksitasi yang disukai, aturan dipilih: di mana r adalah vektor yang permulaannya berada di pusat elemen saraf yang vektornya ditentukan, dan ujungnya berada di tengah elemen saraf ke-n.Aturan dan hukum perubahan biaya dipilih secara empiris, sehingga dapat mensimulasikan pembentukan refleks terkondisi. Lebih detail dalam artikel ini .
mana r adalah vektor yang permulaannya berada di pusat elemen saraf yang vektornya ditentukan, dan ujungnya berada di tengah elemen saraf ke-n.Aturan dan hukum perubahan biaya dipilih secara empiris, sehingga dapat mensimulasikan pembentukan refleks terkondisi. Lebih detail dalam artikel ini . Setelah menerima vektor arah rambatan eksitasi (T) yang lebih disukai, sinapsis (Y1, Y2, Y3 ... Yn) dihitung. Setiap sinaps dicirikan oleh vektor sinaps (S), yang awalnya terletak di pusat neuroelement dan ujungnya dihubungkan dengan pusat neuroelement target di mana sinyal ditransmisikan. Parameter utama sinaps adalah kekuatannya F, nilai gaya dibatasi dalam kerangka kerja tertentu, misalnya, sinaps induksi dapat memiliki nilai dari 0 hingga 10.Bayangkan bahwa vektor T membentuk kerucut di sekitar dirinya yang puncaknya terletak di tengah neuroelement, dan bidang dasar tegak lurus terhadap vektor T, jika vektor sinaps jatuh ke wilayah yang dibatasi oleh kerucut ini, maka nilai sinaps akan meningkat dengan nilai tertentu. Dan karenanya, jika vektor sinaps berada di luar area kerucut, maka gaya sinaps berkurang, tetapi nilai gaya tidak melampaui maksimum dan minimum yang ditetapkan.Wilayah kerucut di sekitar vektor T ditandai dengan sudut pada titik kerucut yang diberikan, sudut ini disebut fokus. Semakin kecil fokus, semakin akurat arah transmisi eksitasi dalam neuroelement akan ditentukan. Seperti yang disebutkan sebelumnya, ketika tubuh mengulangi refleks terkondisi yang sama, itu disempurnakan. Oleh karena itu, metode perubahan fokus berikut dipilih untuk model, ketika menghitung vektor T, itu dibandingkan dengan nilai sebelumnya, dan jika vektor berubah secara tidak signifikan, fokus berkurang dengan nilai tertentu, tetapi jika vektor telah diubah dengan kuat, fokus kembali ke nilai maksimumnya. Hal ini menyebabkan penurunan fokus secara bertahap dengan pengulangan berulang dari kondisi yang sama.Ini adalah poin yang sangat penting, ini adalah seberapa besar kekuatan sinapsis akan berubah dengan setiap aktivasi. Ini ditentukan oleh parameter neuroplastisitas P.Rumus untuk nilai baru gaya sinaps adalah:Fnew = Lipat + I × P × (Fmax - Fmin);Fmin ≥ Fnew ≥ Fmax;di mana P adalah neuroplastisitas (0 ≥ P ≥ 1);I adalah parameter yang menentukan apakah vektor sinapsis berada dalam wilayah peningkatan kekuatan sinapsis (I = 1) atau di wilayah penurunan kekuatan sinapsis (I = -1);Lipat - nilai kekuatan sinapsis sebelumnya;Fmin - nilai minimum gaya sinaps;Fmax - nilai maksimum gaya sinaps.Neuroplastisitas dalam biologi mencirikan bagaimana neuron dapat ditempa terhadap perubahan strukturnya di bawah pengaruh kondisi eksternal. Area otak yang berbeda dicirikan oleh tingkat plastisitasnya sendiri, dan juga dapat bervariasi tergantung pada beberapa faktor.
Setelah menerima vektor arah rambatan eksitasi (T) yang lebih disukai, sinapsis (Y1, Y2, Y3 ... Yn) dihitung. Setiap sinaps dicirikan oleh vektor sinaps (S), yang awalnya terletak di pusat neuroelement dan ujungnya dihubungkan dengan pusat neuroelement target di mana sinyal ditransmisikan. Parameter utama sinaps adalah kekuatannya F, nilai gaya dibatasi dalam kerangka kerja tertentu, misalnya, sinaps induksi dapat memiliki nilai dari 0 hingga 10.Bayangkan bahwa vektor T membentuk kerucut di sekitar dirinya yang puncaknya terletak di tengah neuroelement, dan bidang dasar tegak lurus terhadap vektor T, jika vektor sinaps jatuh ke wilayah yang dibatasi oleh kerucut ini, maka nilai sinaps akan meningkat dengan nilai tertentu. Dan karenanya, jika vektor sinaps berada di luar area kerucut, maka gaya sinaps berkurang, tetapi nilai gaya tidak melampaui maksimum dan minimum yang ditetapkan.Wilayah kerucut di sekitar vektor T ditandai dengan sudut pada titik kerucut yang diberikan, sudut ini disebut fokus. Semakin kecil fokus, semakin akurat arah transmisi eksitasi dalam neuroelement akan ditentukan. Seperti yang disebutkan sebelumnya, ketika tubuh mengulangi refleks terkondisi yang sama, itu disempurnakan. Oleh karena itu, metode perubahan fokus berikut dipilih untuk model, ketika menghitung vektor T, itu dibandingkan dengan nilai sebelumnya, dan jika vektor berubah secara tidak signifikan, fokus berkurang dengan nilai tertentu, tetapi jika vektor telah diubah dengan kuat, fokus kembali ke nilai maksimumnya. Hal ini menyebabkan penurunan fokus secara bertahap dengan pengulangan berulang dari kondisi yang sama.Ini adalah poin yang sangat penting, ini adalah seberapa besar kekuatan sinapsis akan berubah dengan setiap aktivasi. Ini ditentukan oleh parameter neuroplastisitas P.Rumus untuk nilai baru gaya sinaps adalah:Fnew = Lipat + I × P × (Fmax - Fmin);Fmin ≥ Fnew ≥ Fmax;di mana P adalah neuroplastisitas (0 ≥ P ≥ 1);I adalah parameter yang menentukan apakah vektor sinapsis berada dalam wilayah peningkatan kekuatan sinapsis (I = 1) atau di wilayah penurunan kekuatan sinapsis (I = -1);Lipat - nilai kekuatan sinapsis sebelumnya;Fmin - nilai minimum gaya sinaps;Fmax - nilai maksimum gaya sinaps.Neuroplastisitas dalam biologi mencirikan bagaimana neuron dapat ditempa terhadap perubahan strukturnya di bawah pengaruh kondisi eksternal. Area otak yang berbeda dicirikan oleh tingkat plastisitasnya sendiri, dan juga dapat bervariasi tergantung pada beberapa faktor. Contoh ini memungkinkan kita untuk memahami bagaimana refleks terkondisi dibentuk berdasarkan neuroelements asosiatif. Neuroelements putih membentuk busur refleks dari refleks tanpa syarat dengan tajuk "R" dan jawaban "1". Neuroelements ini tidak mengubah nilai kekuatan sinapsisnya. Biru neuroelements awalnya tidak berpartisipasi dalam tindakan refleks, mereka tampaknya mengisi sisa ruang sistem saraf, dan mereka secara tidak sengaja terhubung melalui sinapsis. Oleh karena itu, jika kita mengaktifkan satu elemen saraf yang terkait dengan reseptor "Q", maka fokus tertentu dari kegembiraan akan muncul memiliki sifat acak propagasi dan setelah masuk dalam siklus dengan dirinya sendiri, itu akan keluar setelah beberapa saat, tanpa membuat jawaban apa pun. Jika kita menggabungkan refleks tanpa syarat dengan tajuk "R" dan aktivasi reseptor "Q" pada interval waktu yang kira-kira sama,maka lengkungan refleks dari refleks terkondisi akan terbentuk. Dan sudah aktivasi hanya reseptor "Q" akan mengarah ke jawaban "1".Untuk kejelasan dan optimalisasi model, kami menggunakan penciptaan dinamis elemen saraf, yang mengemulasi ruang penuh sistem saraf dengan elemen yang terhubung secara acak. Di sini, beberapa pertumbuhan neuron baru, atau koneksi baru tidak dimodelkan, semua perubahan hanya terjadi pada kekuatan sinapsis, hanya elemen saraf yang sebelumnya tidak terlibat dalam tindakan refleks tidak ditampilkan.
Contoh ini memungkinkan kita untuk memahami bagaimana refleks terkondisi dibentuk berdasarkan neuroelements asosiatif. Neuroelements putih membentuk busur refleks dari refleks tanpa syarat dengan tajuk "R" dan jawaban "1". Neuroelements ini tidak mengubah nilai kekuatan sinapsisnya. Biru neuroelements awalnya tidak berpartisipasi dalam tindakan refleks, mereka tampaknya mengisi sisa ruang sistem saraf, dan mereka secara tidak sengaja terhubung melalui sinapsis. Oleh karena itu, jika kita mengaktifkan satu elemen saraf yang terkait dengan reseptor "Q", maka fokus tertentu dari kegembiraan akan muncul memiliki sifat acak propagasi dan setelah masuk dalam siklus dengan dirinya sendiri, itu akan keluar setelah beberapa saat, tanpa membuat jawaban apa pun. Jika kita menggabungkan refleks tanpa syarat dengan tajuk "R" dan aktivasi reseptor "Q" pada interval waktu yang kira-kira sama,maka lengkungan refleks dari refleks terkondisi akan terbentuk. Dan sudah aktivasi hanya reseptor "Q" akan mengarah ke jawaban "1".Untuk kejelasan dan optimalisasi model, kami menggunakan penciptaan dinamis elemen saraf, yang mengemulasi ruang penuh sistem saraf dengan elemen yang terhubung secara acak. Di sini, beberapa pertumbuhan neuron baru, atau koneksi baru tidak dimodelkan, semua perubahan hanya terjadi pada kekuatan sinapsis, hanya elemen saraf yang sebelumnya tidak terlibat dalam tindakan refleks tidak ditampilkan. Contoh berikut menunjukkan bagaimana perilaku berperilaku ketika pusat yang berbeda diaktifkan dalam kondisi yang sama dan dengan plastisitas absolut (P = 1).
Contoh berikut menunjukkan bagaimana perilaku berperilaku ketika pusat yang berbeda diaktifkan dalam kondisi yang sama dan dengan plastisitas absolut (P = 1). Perubahan arah propagasi eksitasi di bawah pengaruh dua pusat eksitasi, ketika plastisitas absolut (P = 1):
Perubahan arah propagasi eksitasi di bawah pengaruh dua pusat eksitasi, ketika plastisitas absolut (P = 1):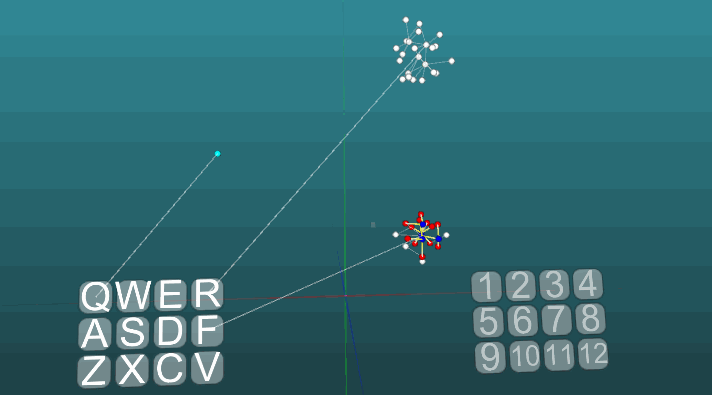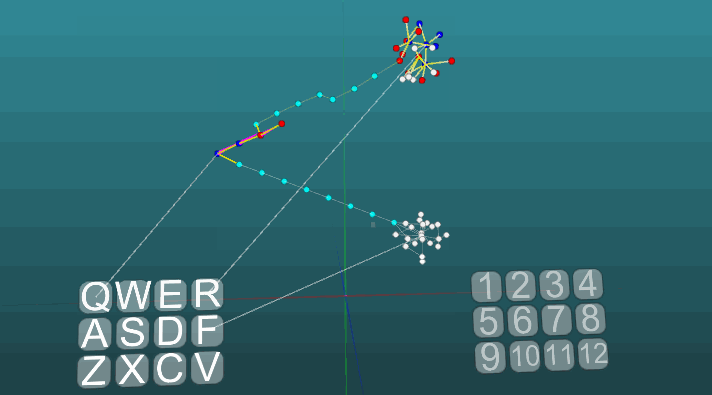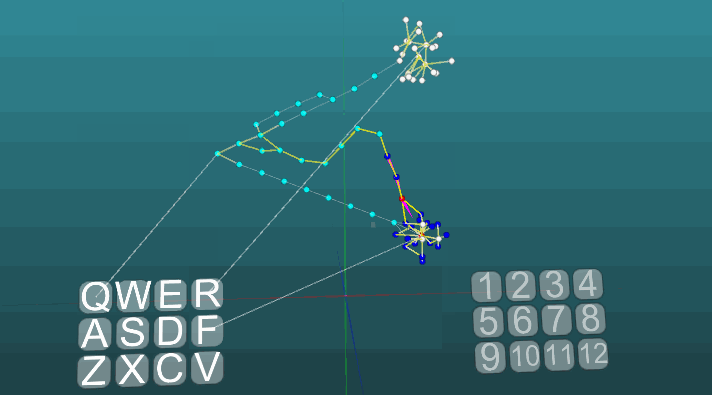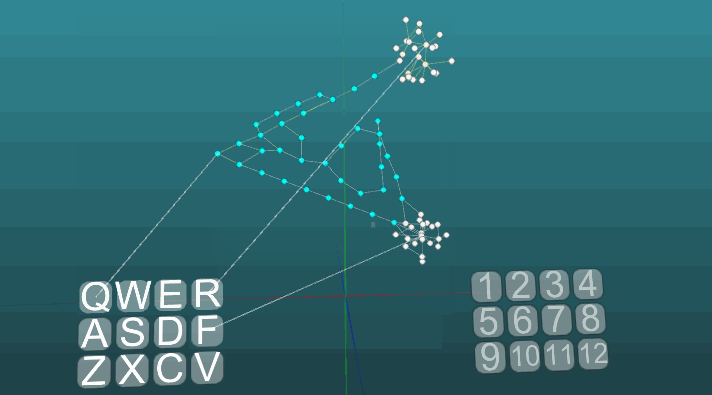 Dan pada plastisitas rendah (P = 0,1):
Dan pada plastisitas rendah (P = 0,1):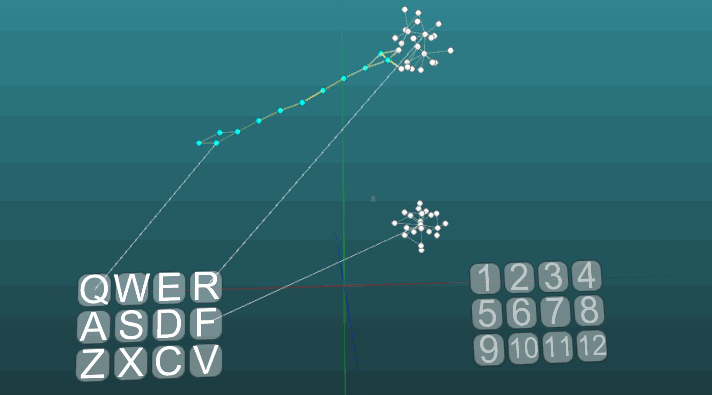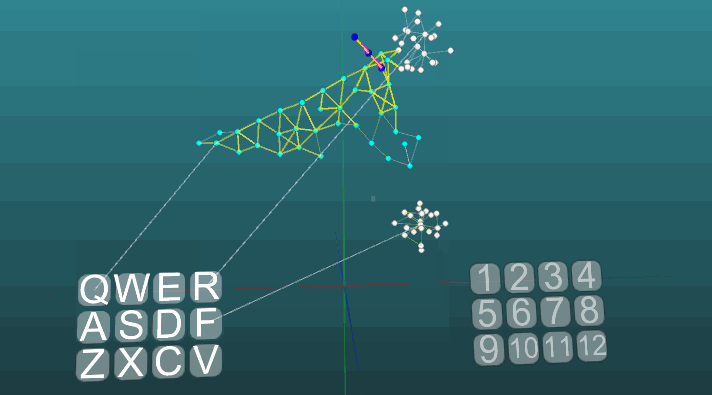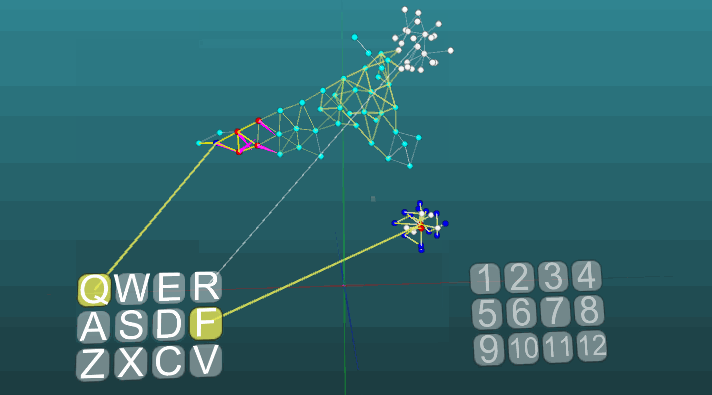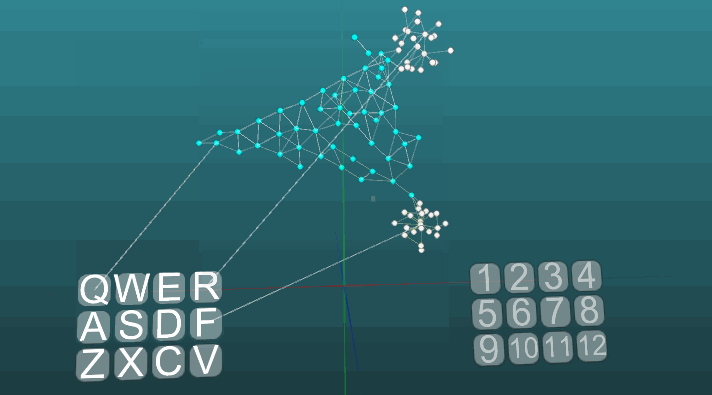 Tentang ini kita telah selesai mempertimbangkan dasar-dasar model sistem saraf. Pada bagian selanjutnya, kita akan mempertimbangkan hal-hal terapan, bagaimana menggunakan semua ini untuk mensimulasikan memori, emosi, dan spesialisasi neuron.
Tentang ini kita telah selesai mempertimbangkan dasar-dasar model sistem saraf. Pada bagian selanjutnya, kita akan mempertimbangkan hal-hal terapan, bagaimana menggunakan semua ini untuk mensimulasikan memori, emosi, dan spesialisasi neuron.