(lanjutan. Bagian sebelumnya: "Otomasi dari DNA dan protein: apa kesamaannya dengan elektronik?" )Kami sepakat pada fakta bahwa operon laktosa mirip dengan elemen AND logis. Tapi dari mana sifat digitalnya berasal? Bagaimanapun, kedua sinyal input (cAMP dan konsentrasi laktosa) sebenarnya adalah analog. Mari kita coba menggambar fungsi input operon laktosa.Setiap pintu masuk dari penekan laktosa terdiri dari dua tahap pengenalan molekul. Laktosa berikatan dengan represor laktosa, dan represor berikatan dengan DNA. Ada beberapa puluh molekul penekan di dalam sel, yang masing-masing, selama tidak ada laktosa, dapat berikatan dengan permulaan operasi laktosa. Ketika terikat di sana, RNA polimerase tidak dapat mulai bekerja. Karena gerakan termal dari molekul penekan, ia terus jatuh dari DNA dan menempel kembali. Jika tidak ada represor sama sekali, operon beroperasi dengan kekuatan penuh (sekarang untuk kesederhanaan kami menganggap bahwa ada kelebihan aktivator katabolik). Pada konsentrasi rendah dari represor, itu mengurangi aktivitas operon hampir linier. Tetapi kemudian efek dari masing-masing bagian baru dari penekan semakin sedikit, dan secara umum, grafik ketergantungan mendekati hiperbola y = 1 / (x +1)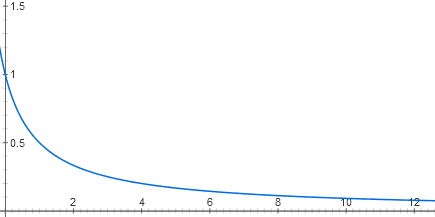
matan. . ?
, , . , :
A + B <-> C
, , - . , - ( , [A], [B] [C] ). : , , .
[C] / ([A] * [B]) = K
. (. . ) . , , .
, , , , ( ) . [C] ([A] * [B]) .
, : RepA + OpA <-> Op ( ). {R-O} = [Op] / ([OpA] * [RepA]). [] ( ) [RepA] ( ). , , : [Op] + [OpA] = 1
[Op] [OpA] :
{R-O} = (1 — [OpA]) / ([OpA] * [RepA])
:
[OpA] = 1 / (K{R-O} * [RepA] + 1)
y = 1 / (x + 1)
Molekul represor mengapung di dalam sel dan berikatan dengan DNA hanya dalam kemasan empat. Agar bentuk represor aktif menjadi tidak aktif, ia harus mengikat empat molekul laktosa - satu atau dua tidak cukup, Anda memerlukan satu molekul laktosa untuk setiap subunit protein dari represor quad. Meskipun ada sedikit laktosa di dalam sel, molekul-molekulnya mengikat satu atau dua penekan dan tetap aktif. Tetapi setelah konsentrasi ambang tertentu laktosa, mayoritas molekul penekan mengikat empat molekul laktosa dan beralih ke bentuk tidak aktif. Grafik aktivitas penekan tergantung pada jumlah laktosa karena itu memiliki bentuk S dan digambarkan oleh fungsi bentuk y = 1 / (1 + x ^ 4).Matan[RepA] [Lac]. , : .
RepA + 4 Lac <-> Rep
:
{R-L} = [Rep] / ([RepA] * [Lac]^4)
, :
[Rep] + [RepA] = n, n ≈ 50
, [RepA] = n / ({R-L} * [Lac]^4 + 1)
Sekarang, untuk mendapatkan karakteristik input operon untuk laktosa, kita harus mengganti fungsi kedua di yang pertama. Ini akan memiliki bentuk, hingga koefisien,y = 1 / (1 + (1 / (1 + x ^ 4))))Jadwalnya juga berbentuk S. Ternyata operon laktosa tidak menanggapi konsentrasi rendah laktosa. Ketika konsentrasi ambang batas tercapai, molekul penekan tidak aktif muncul yang mengikat masing-masing empat laktosa, dan operon menyala agak tajam.Input pengaturan kedua dari operon laktosa juga terdiri dari dua langkah pengenalan molekul: cAMP mengikat aktivator, dan aktivator ke DNA. Perbedaan dari inlet laktosa adalah bahwa di sini pengikatan molekul (aktivator atau cAMP) meningkatkan aktivitas dari apa yang mereka hubungi (operon atau aktivator). Yaitu, grafik akan bergerak dari nol dan mendekati garis horizontal di suatu tempat di atas sumbu X.Matan, , :
2 + Akt <-> AktA
K{-} = [AktA] / ([Akt] * []^2)
, [AktA] [Akt] , 100 , . [Akt] [AktA], :
K{-} = [AktA] / ((100 — [AktA]) * []^2)
[AktA] = K{-} * []^2 / (1 + K{-} * []^2)
[AktA] [] y = x^2 / (1 + x^2)
Karena pengikatan dua molekul cAMP, grafik tidak terlihat seperti hiperbola, tetapi sebagai kurva berbentuk S. Meskipun efek ambang pada itu kurang terlihat daripada pada grafik untuk laktosa, karena derajatnya bukan yang keempat, tetapi hanya yang kedua.Ketergantungan aktivitas operon pada jumlah aktivator memiliki nuansa berbeda. Jika pengikatan represor menekan operon hampir sepenuhnya (di suatu tempat hingga 0,1%, yang lebih rendah dari kesalahan pengukuran biokimia), maka tidak adanya aktivator menekan aktivitas hanya hingga 5%. Oleh karena itu, grafik tidak akan keluar dari nol, tetapi dari titik (0; 0,05):y = 0,05 + (0,95 * x / (1 + x))Matan« , » ,
= [OpA] + 0,05 * [Op]
[OpA] [AktA] . , , [AktA] . , , [AktA] , :
[OpA] = K{-O} * [AktA] / (K{-O} * [AktA] + 1)
y = x / (1 + x)
, ! , (1 — ) * 0,05. , , :
y = 0,05 + (0,95 * x / (1 + x))
Masih untuk mendapatkan ketergantungan dari aktivitas operon pada konsentrasi cAMP. Untuk melakukan ini, gantikan formula pertama (untuk konsentrasi CAP aktif tergantung pada konsentrasi cAMP) dalam formula kedua (untuk aktivitas operon dari konsentrasi ATS aktif) ...... Dan dapatkan fraksi empat lantai:Kami mendapat fungsi paling kompleks dari semuanya di artikel ini. Namun, grafiknya terlihat seperti grafik ketergantungan sederhana dari CAP aktif pada konsentrasi cAMP, y = x ^ 2 / (1 + x ^ 2):Ini juga merupakan kurva berbentuk S dengan beberapa efek ambang, yang perlahan-lahan mendekati horizontal y = 1. Itu hanya dimulai bukan dari nol, tetapi dari titik (0; 0,05).Masih memahami bagaimana kedua input berinteraksi. Dalam kasus operon laktosa, jawabannya sederhana - tidak mungkin. Penekan laktosa dan aktivator katabolik tidak mempengaruhi pengikatan satu sama lain dengan DNA. Oleh karena itu, pengikatan kedua protein ini dapat dianggap peristiwa independen. Aktivitas penuh operon dicapai ketika kedua aktivator terhubung dan represor tidak terhubung. Peluang kebetulan seperti itu sama dengan produk probabilitas masing-masing secara terpisah. Jadi, untuk mendapatkan fungsi aktivitas operon laktosa dari dua variabel ([Lac]) dan ([cAMP]), Anda hanya perlu mengalikan fungsi dari masing-masing variabel ini:Aktivitas = (1 / (1 + (1 / (1 + [Lac] ^ 4)))) * (0,05 + 0,095 * ([cAMP] ^ 2 / (1 + [cAMP] ^ 2)) / ( 1 + ([cAMP] ^ 2 / (1 + [cAMP] ^ 2)))))Grafik tiga dimensi dari fungsi iniItu tampak seperti dataran tinggi yang berakhir di ngarai di dua sisi. Ngarai kanan (konsentrasi laktosa rendah) lebih dalam dan lebih datar dari kiri (konsentrasi cAMP rendah).Dalam kehidupan nyata, E. coli di sekitarnya tidak ada laktosa sama sekali (paling sering), atau konsentrasinya lebih tinggi dari ambang batas dan penekan laktosa praktis tidak mengganggu operasi operon (ketika pemilik makan sesuatu susu). Konsentrasi cAMP adalah sinyal internal yang diproduksi oleh sel itu sendiri. Juga terlalu kecil untuk memasukkan operon laktosa (ketika ada glukosa atau pati), atau cukup untuk menyalakan 95% atau lebih (jika tidak ada yang lebih enak daripada laktosa). Artinya, hampir selalu operon laktosa berada dalam kondisi dataran tinggi pada grafik ini, atau di salah satu ngarai.Fungsi input operon laktosa diukur secara eksperimental. Metode pengukuran menyerupai teknik debugging mikrokontroler "menggantung LED pada kaki output." Wilayah pengaturan operon diambil dan dihubungkan dengan gen protein fluorescent hijau. Konstruk gen ini dimasukkan ke dalam sel-sel Escherichia coli, dari mana operon laktosa reguler dihapus (sehingga konsentrasi laktosa diatur secara ketat oleh eksperimen) dan sistem pengkodean standar kelaparan dengan tingkat cAMP. Setelah itu, aktivitas operon laktosa dapat diukur secara akurat dengan spektrofotometer sesuai dengan tingkat fluoresensi hijau. Ternyata, dalam batas keakuratan pengukuran (1%), teori sepenuhnya menyatu dengan eksperimen ).Bagian bawah ngarai kiri pada grafik fungsi input (pada konsentrasi rendah cAMP) di atas nol. Ini bukan bug, tetapi fitur: berkat itu, di hadapan beberapa gula sekaligus, E. coli siap untuk dengan cepat beralih ke laktosa ketika glukosa berakhir. Escherichia coli mutan, di mana fungsi pengaturan operon laktosa lebih dekat dengan murni DAN (yaitu, operon tidak bekerja tanpa cAMP), beralih dari glukosa ke laktosa selama satu jam atau lebih: pada saat glukosa habis, mereka tidak memiliki enzim penyerapan laktosa, yang berarti bahwa tidak ada enzim serapan laktosa, yang berarti bahwa tidak ada enzim serapan laktosa, energi untuk membuatnya dengan cepat. Escherichia coli normal ("tipe liar") memenuhi titik ini, sudah memiliki beberapa enzim untuk penyerapan laktosa dan dibutuhkan 15-20 menit untuk sepenuhnya beralih ke gula baru.Sekarang mari kita menyimpang dari operon laktosa dan melihat lebih luas pada apa elemen logis pada suatu basis kimia mampu. Hampir di mana-mana dalam sistem pensinyalan biokimia kita menemukan pengenalan molekuler, yang digambarkan oleh fungsi bentuk y = x ^ n / (1 + x ^ n) untuk aktivator dan y = 1 / (1 + x ^ n) untuk represor. Tingkat n dalam fungsi-fungsi ini menunjukkan jumlah molekul dari jenis yang sama untuk mengikat untuk mendapatkan efek, dan biasanya sama dengan 1, 2, atau 4 (paling sering 2). Kunci logis yang lebih kompleks dapat diperoleh dengan menambahkan lebih banyak situs pengikat protein pengatur ke awal operon. Mungkin ada lebih dari dua jenis protein ini. Efek dari berbagai protein pada pendaratan RNA polimerase dapat bertambah (OR) atau bertambah banyak (DAN).Tetapi secara umum, kita akan berurusan dengan beberapa kombinasi (penambahan atau perkalian) dari fungsi-fungsi dasar y = x ^ n / (1 + x ^ n) dan y = 1 / (1 + x ^ n) dengan derajat dan koefisien yang berbeda.Beberapa fitur menarik sangat sederhana. Misalnya, protein pengatur yang sama dapat mengikat (dalam bentuk berpasangan) ke dua wilayah promotor. Di satu bagian, itu bertindak sebagai penekan, dan di bagian lain - sebagai aktivator. Agar operon berfungsi, situs represor harus kosong, dan situs aktivator sibuk. Ternyata fungsi dengan maksimal jelas:y = (1 / (1 + x ^ 2)) * (x ^ 2 / (1 + x ^ 2))Tanpa protein ini, operon tidak aktif karena situs aktivator kosong, dan pada konsentrasi tinggi tidak aktif karena situs sibuk penekan. Aktivitas maksimum akan terjadi ketika situs aktivator dan situs represor ditempati dengan probabilitas 50%.Anda masih dapat membuat dua situs pengikat aktivator, di satu aktivator akan mengikat kuat pada DNA, tetapi mengaktifkan gen dengan lemah, di lain - lemah menempel pada DNA, tetapi sangat mengaktifkan gen. Dalam kasus ini, kita mendapatkan fungsi seperti y = x ^ 4 / (1 + x ^ 4) + 0.3 * (10x) ^ 4 / (1 + (10x) ^ 4), dengan grafik bertahap:Matematika, ay! Bisakah Anda memberi tahu di komentar apa lagi yang mungkin dan apa yang tidak dapat diperoleh dengan menambahkan dan mengalikan fungsi dasar ini y = x ^ n / (1 + x ^ n) dan y = 1 / (1 + x ^ n)?Pada bagian selanjutnya, kita akan berurusan dengan rangkaian elemen logis semacam itu.