← Artikel sebelumnya dalam siklusLogika untuk memasukkan operon dari jalur metabolisme yang berbeda di E. coli dekat dengan elemen AND dan OR. Oleh karena itu, berdasarkan pada mereka, Anda dapat membuat konstruksi gen yang mengimplementasikan skema logis arbitrer (tidak terlalu rumit). Ada beberapa program, misalnya, Cello, yang, dari uraian logika di Verilog, membuat urutan konstruk gen yang mengimplementasikan logika ini.Masalah terbalik ternyata lebih sulit: untuk mendapatkan deskripsi logika pengaturannya dari urutan DNA alami. Mari kita mulai dengan kasus-kasus sederhana di mana tidak ada loop umpan balik yang kompleks dan integrasi banyak sinyal pada satu operon. Hampir semua regulasi gen E. coli memenuhi kondisi ini. Jika kita mengambil semua koneksi pengaturan yang diketahui antara gen Escherichia coli dan membangun jaringannya, maka kita dapat menghitung statistik "motif" jaringan ini (metode elemen penghubung) dan memilih yang paling sering digunakan.Gene Switching Dynamics
Pada artikel terakhir, kami memodelkan operasi operon laktosa dengan model sederhana dengan empat parameter. Waktu bukan bagian dari model ini. Oleh karena itu, pada kenyataannya, hanya menggambarkan keadaan keseimbangan, ketika level sinyal input tetap konstan untuk waktu yang lama. Untuk memodelkan proses cepat ketika menghidupkan dan mematikan gen, faktor-faktor tambahan harus dipertimbangkan.Pertama, ada penundaan transkripsi: dari awal kerja RNA polimerase ke akhir sintesis mRNA, dan kemudian protein, waktu yang nyata berlalu, sekitar 10 menit dalam Escherichia coli dan dari satu jam atau lebih pada manusia. Oleh karena itu, tingkat penampilan molekul protein baru tidak tergantung pada keadaan promotor gennya sekarang, tetapi pada keadaan promotor beberapa menit yang lalu.Kedua, setiap protein tidak hanya disintesis, tetapi juga dipecah. Dalam kasus yang paling sederhana, pemecahan protein tidak terkontrol dan setiap menit sebagian kecil molekul protein ini terurai dalam sel. Semakin banyak mereka, semakin banyak mereka putus. Artinya, protein memiliki waktu paruh, di mana konsentrasinya turun hingga setengahnya.Lebih detail untuk yang paling penasaran— - . , — .
Untuk menggambarkan dinamika menghidupkan dan mematikan gen, kita perlu menulis sebuah diffur:d[]dt=A(t−tau)–k∗[]
di mana A adalah aktivitas gen (fungsi input yang sama dari artikel sebelumnya), tau adalah waktu tunda transkripsi, dan k adalah tingkat pemecahan protein. Jika A pertama sama dengan nol, dan kemudian naik tiba-tiba dan tetap pada tingkat yang konstan, maka perubahan jumlah protein dari waktu ke waktu akan terlihat seperti ini:Motif dalam jaringan gen dan fungsinya
Karakteristik motif paling sederhana dari Escherichia coli - loop umpan balik - terdiri dari satu gen yang mengatur dirinya sendiri. Paling sering, gen adalah penekan untuk dirinya sendiri, yaitu, umpan balik dalam loop negatif. Jika ini adalah input pengaturan utama, maka umpan balik ini biasanya berfungsi untuk menstabilkan tingkat protein yang dikodekan oleh gen. Loop umpan balik semacam itu, misalnya, mempertahankan tingkat penekan laktosa yang konstan dalam sel. Jika, selain umpan balik negatif, ada input pengaturan lainnya, maka umpan balik mempercepat respons gen terhadap sinyal eksternal.(garis putus-putus adalah kurva teoritis, garis padat adalah hasil percobaan. Garis hijau dengan umpan balik negatif, garis biru tanpa itu)Umpan balik positif (protein mengaktifkan gennya sendiri) biasanya mengarah pada perilaku pemicu. Gen semacam itu memiliki dua keadaan stabil: tidak berfungsi sama sekali, atau bekerja dengan kapasitas penuh. Sinyal eksternal yang kuat tetapi pendek dapat mentransfer gen dari satu keadaan stabil ke yang lain di mana ia tetap sampai sinyal berikutnya. Motif semacam itu dapat berfungsi sebagai elemen memori. Jika suatu gen “menangkap” noise pada input regulatori, maka umpan balik positif akan menyebabkan penyebaran yang kuat dalam aktivitas gen antara sel-sel tetangga yang tumbuh di bawah kondisi yang sama. Ini digunakan untuk menjaga heterogenitas bakteri yang tumbuh bersama sehingga stres mendadak tidak membunuh semua orang sekaligus.Sebagai contoh, banyak antibiotik mengganggu sintesis protein. Jika sel tidak tumbuh dan tidak mensintesis protein, maka ia dapat dengan mudah bertahan dari penambahan antibiotik ke medium dan kembali tumbuh nanti. Pada E. coli, bahkan dalam kondisi ideal, hingga 0,01% sel berada dalam hibernasi jika terjadi keracunan tiba-tiba pada lingkungan. Sel-sel aktif ini (mereka disebut "persistors") tidak berbeda dari yang lain dalam DNA, mereka masuk dan keluar dari hibernasi di bawah kendali gen dengan umpan balik positif dan input noise.Motif umum lainnya dari jaringan gen sedikit lebih rumit. Ini disebut "umpan maju" dan terdiri dari tiga gen: gen X mengatur gen Y dan Z, gen Y mengatur Z. Masing-masing dari tiga tautan dalam loop dapat aktivasi atau represi, oleh karena itu, pada prinsipnya, 8 jenis dimungkinkan loop seperti itu. Pada kenyataannya, biasanya dua tipe delapan ditemukan: gen X dan Y adalah aktivator (loop koheren tipe 1), atau X adalah aktivator Y dan Z, Y adalah represor Z (loop 1 koheren tipe).Perilaku loop dengan dua aktivator tergantung pada logika interaksi input pada Z. Jika gen output bekerja sebagai AND dalam loop, maka loop bekerja seperti filter low-pass yang tidak melewatkan pulsa pendek ke output. Agar sinyal output muncul dalam loop seperti itu, input (X) harus menerima sinyal yang cukup lama untuk protein terakumulasi, dan kemudian protein Y. Hanya aksi gabungan dari protein X dan Y yang akan mengaktifkan gen Z.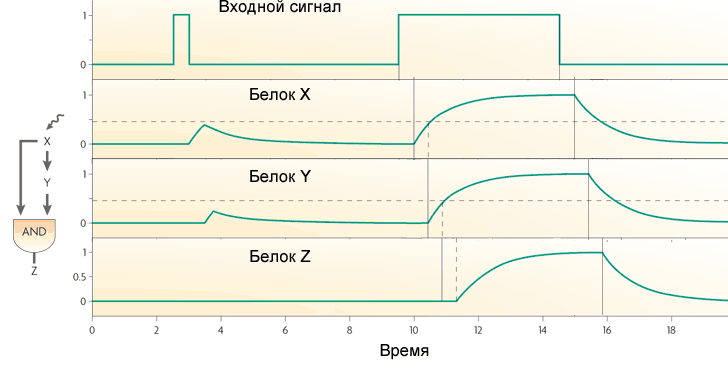 Jika gen Z bekerja seperti OR, maka fungsi loop menjadi kebalikannya: ia meneruskan semua pulsa ke output, dan berubah pendek menjadi panjang. Tapi dia tidak melewatkan jeda singkat di antara pulsa.Perulangan langsung dengan aktivator X dan represor Y bertindak sebagai filter lintasan tinggi. Ini menghasilkan pulsa dengan panjang tetap kecil (sama dengan keterlambatan transkripsi gen Y) pada output sebagai respons terhadap tepi terdepan dari sinyal input.Motif ketiga yang sering dalam jaringan gen Escherichia coli adalah modul input tunggal. Ini hanyalah sekelompok gen yang digerakkan oleh gen pengatur yang sama. Biasanya, gen dalam kelompok ini memiliki ambang aktivasi yang berbeda, sehingga peningkatan yang lancar dalam konsentrasi faktor transkripsi mencakup gen dalam urutan tertentu. Skema semacam itu digunakan, misalnya, dalam mengelola respons terhadap stres.
Jika gen Z bekerja seperti OR, maka fungsi loop menjadi kebalikannya: ia meneruskan semua pulsa ke output, dan berubah pendek menjadi panjang. Tapi dia tidak melewatkan jeda singkat di antara pulsa.Perulangan langsung dengan aktivator X dan represor Y bertindak sebagai filter lintasan tinggi. Ini menghasilkan pulsa dengan panjang tetap kecil (sama dengan keterlambatan transkripsi gen Y) pada output sebagai respons terhadap tepi terdepan dari sinyal input.Motif ketiga yang sering dalam jaringan gen Escherichia coli adalah modul input tunggal. Ini hanyalah sekelompok gen yang digerakkan oleh gen pengatur yang sama. Biasanya, gen dalam kelompok ini memiliki ambang aktivasi yang berbeda, sehingga peningkatan yang lancar dalam konsentrasi faktor transkripsi mencakup gen dalam urutan tertentu. Skema semacam itu digunakan, misalnya, dalam mengelola respons terhadap stres.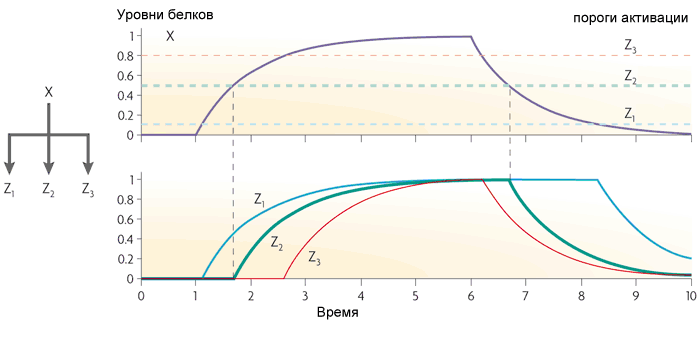 Motif keempat, yang paling kompleks adalah modul multi-input, atau regulator yang tumpang tindih padat. Ini terdiri dari beberapa gen pengatur yang mengendalikan beberapa gen "output".Bergantung pada logika pengaturan setiap gen keluaran, motif ini dapat bekerja sangat berbeda. Ini dapat dibandingkan dengan kumpulan elemen logika, yang, tergantung pada koneksi, dapat berupa dekoder, penghitung, atau yang lainnya. Deskripsi lengkap tentang logika karya motif semacam itu belum ada.Dengan keempat motif ini, struktur jaringan gen Escherichia coli atau ragi praktis habis. Atas dasar mereka, berbagai skema buatan sudah dibuat. Misalnya, bakteri dengan generator osilasi, yang juga menyinkronkan dalam sel tetangga, dan output protein luminescent
Motif keempat, yang paling kompleks adalah modul multi-input, atau regulator yang tumpang tindih padat. Ini terdiri dari beberapa gen pengatur yang mengendalikan beberapa gen "output".Bergantung pada logika pengaturan setiap gen keluaran, motif ini dapat bekerja sangat berbeda. Ini dapat dibandingkan dengan kumpulan elemen logika, yang, tergantung pada koneksi, dapat berupa dekoder, penghitung, atau yang lainnya. Deskripsi lengkap tentang logika karya motif semacam itu belum ada.Dengan keempat motif ini, struktur jaringan gen Escherichia coli atau ragi praktis habis. Atas dasar mereka, berbagai skema buatan sudah dibuat. Misalnya, bakteri dengan generator osilasi, yang juga menyinkronkan dalam sel tetangga, dan output protein luminescent — « » «Hello World» . « » ( , ), , , , , , (
).
. . , .