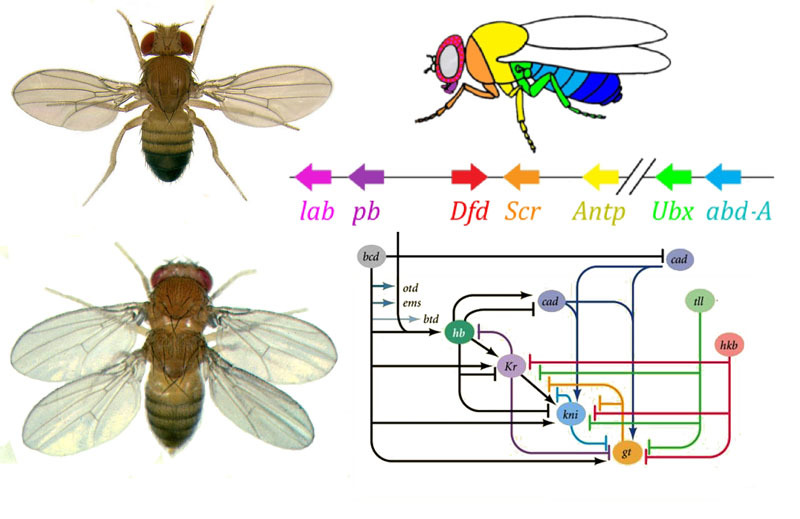
 Bagaimana cara menggunakan telur untuk mengubah telur menjadi larva dan kemudian menjadi lalat dewasa? Seekor lalat, seperti serangga lainnya, terdiri dari segmen-segmen. Segmen digabungkan dalam tiga kelompok - kepala, dada dan perut. Segmen mungkin memiliki sepasang anggota badan. Ada tiga segmen di dada, masing-masing dengan sepasang kaki. Segmen toraks kedua juga memiliki sayap, dan yang ketiga - kumbang darat (sayap ekor kecil, hampir tidak terlihat tanpa kaca pembesar). Perut terdiri dari delapan segmen, tidak ada anggota badan pada mereka. Awalnya, ada enam segmen di kepala, tetapi selama evolusi serangga, batas-batas di antara mereka benar-benar terhapus. Tungkai mereka adalah antena yang dengannya lalat mengendus dan merasakan, dan tiga pasang anggota gerak mulut. Untuk bug atau kecoak, itu adalah rahang bawah, rahang atas dan bibir bawah, sedangkan untuk lalat mereka bergabung ke dalam belalai.Agar semua organ berada di tempatnya, setiap sel harus tahu di mana letaknya di dalam embrio. Oleh karena itu, pada tahap awal perkembangan embrio, ketika belum ada organ di dalamnya dan semua sel terlihat sama, sebuah “kotak koordinat” biokimia muncul di dalamnya dengan sumbu anteroposterior, dorso-abdomen, dan kiri-kanan. Mutasi gen yang membentuk grid koordinat dapat dengan mudah dan cepat mengubah penampilan hewan - memindahkan organ ke tempat lain, mengubah jumlah mereka, dan sebagainya. Oleh karena itu, gen semacam itu penting untuk evolusi hewan dan sangat aktif dipelajari. Jaringan gen untuk menandai embrio lalat Drosophila, mainan genetika favorit, mungkin yang paling banyak dipelajari di antara jaringan gen hewan.Algoritma untuk menandai telur di sepanjang sumbu anteroposterior pada dasarnya sederhana:1) menentukan ujung telur yang akan menjadi kepala, yang ekornya2) membagi embrio sepanjang sumbu anteroposterior menjadi 17 segmen3) membagi setiap segmen ke depan, tengah dan belakang4) tandai kepala (6 segmen), dada (3 segmen) dan perut (8 segmen)5) tandai perbedaan individu dari segmen-segmen Jaringangen yang berurusan dengan ini memiliki perangkat modular. Setiap modul melakukan langkahnya sendiri dari algoritma umum. Lalat Drosophila ternyata menjadi objek yang sangat nyaman untuk mempelajari jaringan pengembangan gen, karena memiliki kelainan bawaan yang sangat cerah dan mengungkapkan. Misalnya, mutasi antennapedia di mana antena diubah menjadi kaki atau
Bagaimana cara menggunakan telur untuk mengubah telur menjadi larva dan kemudian menjadi lalat dewasa? Seekor lalat, seperti serangga lainnya, terdiri dari segmen-segmen. Segmen digabungkan dalam tiga kelompok - kepala, dada dan perut. Segmen mungkin memiliki sepasang anggota badan. Ada tiga segmen di dada, masing-masing dengan sepasang kaki. Segmen toraks kedua juga memiliki sayap, dan yang ketiga - kumbang darat (sayap ekor kecil, hampir tidak terlihat tanpa kaca pembesar). Perut terdiri dari delapan segmen, tidak ada anggota badan pada mereka. Awalnya, ada enam segmen di kepala, tetapi selama evolusi serangga, batas-batas di antara mereka benar-benar terhapus. Tungkai mereka adalah antena yang dengannya lalat mengendus dan merasakan, dan tiga pasang anggota gerak mulut. Untuk bug atau kecoak, itu adalah rahang bawah, rahang atas dan bibir bawah, sedangkan untuk lalat mereka bergabung ke dalam belalai.Agar semua organ berada di tempatnya, setiap sel harus tahu di mana letaknya di dalam embrio. Oleh karena itu, pada tahap awal perkembangan embrio, ketika belum ada organ di dalamnya dan semua sel terlihat sama, sebuah “kotak koordinat” biokimia muncul di dalamnya dengan sumbu anteroposterior, dorso-abdomen, dan kiri-kanan. Mutasi gen yang membentuk grid koordinat dapat dengan mudah dan cepat mengubah penampilan hewan - memindahkan organ ke tempat lain, mengubah jumlah mereka, dan sebagainya. Oleh karena itu, gen semacam itu penting untuk evolusi hewan dan sangat aktif dipelajari. Jaringan gen untuk menandai embrio lalat Drosophila, mainan genetika favorit, mungkin yang paling banyak dipelajari di antara jaringan gen hewan.Algoritma untuk menandai telur di sepanjang sumbu anteroposterior pada dasarnya sederhana:1) menentukan ujung telur yang akan menjadi kepala, yang ekornya2) membagi embrio sepanjang sumbu anteroposterior menjadi 17 segmen3) membagi setiap segmen ke depan, tengah dan belakang4) tandai kepala (6 segmen), dada (3 segmen) dan perut (8 segmen)5) tandai perbedaan individu dari segmen-segmen Jaringangen yang berurusan dengan ini memiliki perangkat modular. Setiap modul melakukan langkahnya sendiri dari algoritma umum. Lalat Drosophila ternyata menjadi objek yang sangat nyaman untuk mempelajari jaringan pengembangan gen, karena memiliki kelainan bawaan yang sangat cerah dan mengungkapkan. Misalnya, mutasi antennapedia di mana antena diubah menjadi kaki atau VROTMNENOGIproboscipedia, ketika bagian dari belalai berubah menjadi kaki. Mutasi bithorax memberikan lalat bersayap empat, yang, di samping sayap yang biasa pada segmen toraks kedua, memiliki sepasang sayap kedua pada segmen toraks ketiga.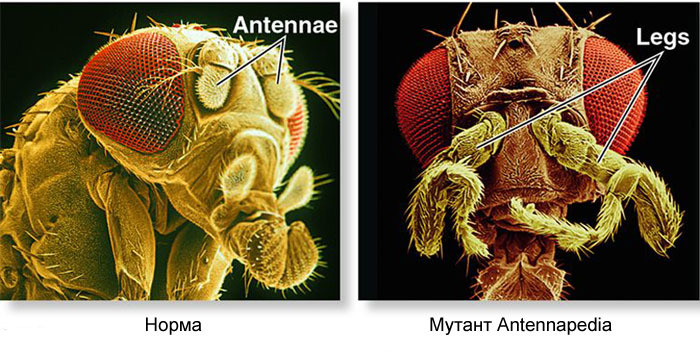
 Mutasi seperti itu, di mana satu bagian tubuh menjadi sangat mirip dengan yang lain, disebut "homeosis". Dimulai dengan mutan ini, genetika akhirnya menguraikan seluruh jaringan gen yang mengendalikan perkembangan lalat. Karya ini memenangkan Hadiah Nobel pada tahun 1995.Semuanya dimulai dengan gen polaritas telur - bicoid dan nano. Mereka bekerja bahkan sebelum pembuahan sel telur, sementara itu matang di ovarium lalat induk. Sel bantu, memompa nutrisi ke dalam telur, terletak di satu sisi telur, sehingga telur memiliki dua ujung yang berbeda dari awal. Matriks bicoid RNA terakumulasi di ujung anterior telur, di mana ia melekat dan tidak bisa berenang menjauh. nanos mRNA juga difiksasi pada ujung posterior. Setelah pembuahan, protein mulai disintesis menggunakan mRNA ini. Molekul mereka menyebar ke seluruh sel telur. Karena sintesis protein bicoid dan nanos terjadi pada satu ujung telur, dan membusuk di mana-mana, gradien protein bicoid berkurang, menurun dari ujung anterior embrio ke posterior, dan gradien nanos dari posterior ke ujung anterior.
Mutasi seperti itu, di mana satu bagian tubuh menjadi sangat mirip dengan yang lain, disebut "homeosis". Dimulai dengan mutan ini, genetika akhirnya menguraikan seluruh jaringan gen yang mengendalikan perkembangan lalat. Karya ini memenangkan Hadiah Nobel pada tahun 1995.Semuanya dimulai dengan gen polaritas telur - bicoid dan nano. Mereka bekerja bahkan sebelum pembuahan sel telur, sementara itu matang di ovarium lalat induk. Sel bantu, memompa nutrisi ke dalam telur, terletak di satu sisi telur, sehingga telur memiliki dua ujung yang berbeda dari awal. Matriks bicoid RNA terakumulasi di ujung anterior telur, di mana ia melekat dan tidak bisa berenang menjauh. nanos mRNA juga difiksasi pada ujung posterior. Setelah pembuahan, protein mulai disintesis menggunakan mRNA ini. Molekul mereka menyebar ke seluruh sel telur. Karena sintesis protein bicoid dan nanos terjadi pada satu ujung telur, dan membusuk di mana-mana, gradien protein bicoid berkurang, menurun dari ujung anterior embrio ke posterior, dan gradien nanos dari posterior ke ujung anterior.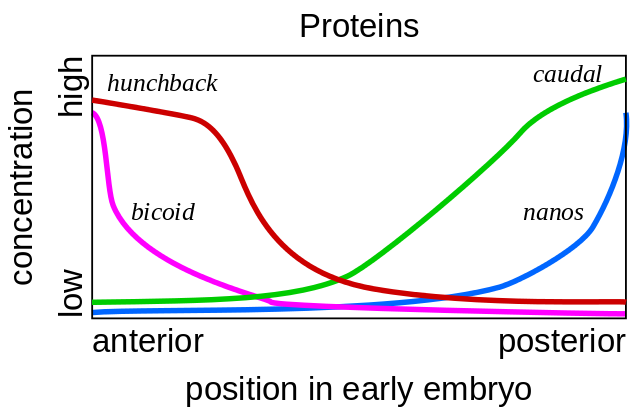 Jika mRNA bicoid dimasukkan ke tempat yang berbeda dalam telur dengan mikropipet, kepala kedua akan tumbuh di sana. Jika mRNA bicoid secara artifisial dimatikan, embrio diperoleh tanpa kepala dan dengan dua keledai.Lebih lanjut, protein bicoid dan nanos meliputi gen embrio itu sendiri, yaitu, mRNA baru mulai dibaca dari mereka). Gen gap adalah yang pertama mulai bekerja, dari gap bahasa Inggris, gap. Dua gen pertama dari kelompok ini - bongkok dan ekor - membentuk gradien dari kepala ke ekor (bongkok) dan dari ekor ke kepala (ekor). Tapi gradien mereka lebih halus daripada bicoid dan nano.
Jika mRNA bicoid dimasukkan ke tempat yang berbeda dalam telur dengan mikropipet, kepala kedua akan tumbuh di sana. Jika mRNA bicoid secara artifisial dimatikan, embrio diperoleh tanpa kepala dan dengan dua keledai.Lebih lanjut, protein bicoid dan nanos meliputi gen embrio itu sendiri, yaitu, mRNA baru mulai dibaca dari mereka). Gen gap adalah yang pertama mulai bekerja, dari gap bahasa Inggris, gap. Dua gen pertama dari kelompok ini - bongkok dan ekor - membentuk gradien dari kepala ke ekor (bongkok) dan dari ekor ke kepala (ekor). Tapi gradien mereka lebih halus daripada bicoid dan nano.Mengapa gen begitu anehnya disebut?. . , hunchback — «», tailless — «», kruppel - — «», bithorax – « ». Sonic hedgehog , .
Empat gen yang tersisa dari kelompok gap masing-masing membentuk satu atau dua pita lebar (lebih dari satu segmen) dengan batas yang cukup jelas. Mereka digabungkan oleh produk protein dari gen yang lebih tinggi (bicoid, nanos, bongkok dan ekor). Pematian total salah satu gen dalam kelompok ini memberikan embrio yang dipersingkat, yang tidak memiliki beberapa segmen, yaitu gen-gen di mana gen ini biasanya bekerja.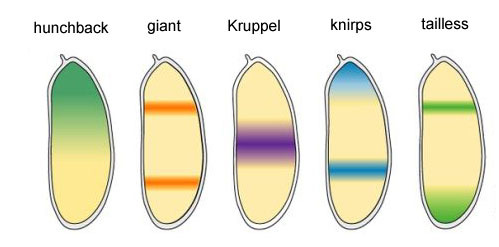 Area aktivitas gen blok celahMengapa ternyata keempat gen blok celah (kruppel, raksasa, knirps, dan berekor) membagi embrio menjadi pita, di mana masing-masing dari satu gen itu bekerja?Pertama, mereka semua menekan aktivitas masing-masing. Dalam satu sel, suatu kondisi akan stabil di mana satu gen dari kelompok seperti itu secara aktif bekerja dan menekan yang lainnya (pemicu). Tetapi telur lalat yang berkembang lebih kompleks. Di dalamnya, pada saat ini, ribuan inti sel individu mengapung dalam cairan intraseluler total (sitoplasma). Ukuran telur cukup besar sehingga laju pembusukan protein pengatur sebanding dengan tingkat difusi mereka ke ujung telur lainnya. Dengan kombinasi difusi dan saling menekan, keadaan stabil lainnya muncul ketika telur dibagi menjadi beberapa pita, di mana masing-masing satu gen dari kelompok bekerja. Semakin lambat difusi protein atau semakin cepat penguraiannya, semakin kecil garis-garis atau bintik-bintik ini.Partisipasi dari sistem tersebut (mereka disebut reaksi-difusi) dalam penandaan tubuh hewan diprediksi oleh Alan Turing (ya, yang menemukan "mesin Turing") pada tahun 1952. Pada waktu itu, sedikit yang diketahui tentang gen dan protein, tetapi persamaan yang menggambarkan struktur bergaris-garis dan berbintik itu disimpulkan dengan benar oleh Turing.Di sini atau di sini, Anda dapat bermain dengan model difusi reaksi sederhana dari dua zat dan merasakan bagaimana ia dapat berperilaku.
Area aktivitas gen blok celahMengapa ternyata keempat gen blok celah (kruppel, raksasa, knirps, dan berekor) membagi embrio menjadi pita, di mana masing-masing dari satu gen itu bekerja?Pertama, mereka semua menekan aktivitas masing-masing. Dalam satu sel, suatu kondisi akan stabil di mana satu gen dari kelompok seperti itu secara aktif bekerja dan menekan yang lainnya (pemicu). Tetapi telur lalat yang berkembang lebih kompleks. Di dalamnya, pada saat ini, ribuan inti sel individu mengapung dalam cairan intraseluler total (sitoplasma). Ukuran telur cukup besar sehingga laju pembusukan protein pengatur sebanding dengan tingkat difusi mereka ke ujung telur lainnya. Dengan kombinasi difusi dan saling menekan, keadaan stabil lainnya muncul ketika telur dibagi menjadi beberapa pita, di mana masing-masing satu gen dari kelompok bekerja. Semakin lambat difusi protein atau semakin cepat penguraiannya, semakin kecil garis-garis atau bintik-bintik ini.Partisipasi dari sistem tersebut (mereka disebut reaksi-difusi) dalam penandaan tubuh hewan diprediksi oleh Alan Turing (ya, yang menemukan "mesin Turing") pada tahun 1952. Pada waktu itu, sedikit yang diketahui tentang gen dan protein, tetapi persamaan yang menggambarkan struktur bergaris-garis dan berbintik itu disimpulkan dengan benar oleh Turing.Di sini atau di sini, Anda dapat bermain dengan model difusi reaksi sederhana dari dua zat dan merasakan bagaimana ia dapat berperilaku.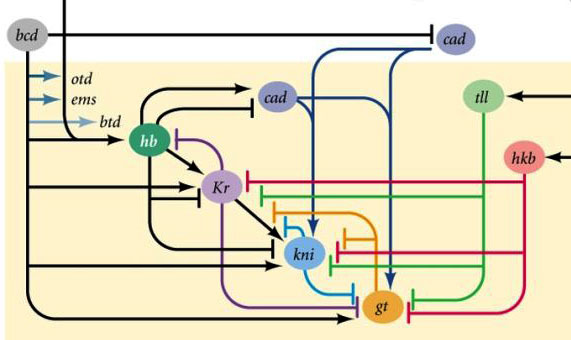 Hubungan antara gen gap dan gen polaritas telur. Bcd - bicoid, hb - bongkok, cad - caudal, Kr - Kruppel, kni - knirps, gt - giant, tll - berekor.Penindasan gen masing-masing saja akan menandai embrio menjadi beberapa garis, tetapi lokasi pita aktivitas dari empat gen relatif satu sama lain dan ujung embrio akan sangat bervariasi. Agar strip selalu ada di tempat (misalnya, untuk knirps - satu strip di kepala, yang lain di bagian belakang antara kruppel dan strip raksasa), Anda memerlukan sinyal kontrol kedua - gradien bicoid dan protein polaritas telur lainnya. Fungsi input gen gap untuk regulator ini memiliki maksimum pada berbagai level sinyal input (kami memperoleh fungsi input seperti itu di akhir artikel kedua ). Oleh karena itu, gen-gen celah masing-masing dihidupkan oleh tingkat protein bicoid, hunchback, dan caudal mereka, dan zona aktivitas mereka secara alami terletak dari kepala ke ekor embrio.Setelah gen gap membuat tanda kasar embrio menjadi 7 pita (mereka tidak bertepatan dengan segmen atau kepala-dada-perut), modul berikutnya dihidupkan - aturan pasangan ("aturan aturan pasangan"). Itu sudah membagi embrio menjadi segmen, yang seharusnya 17. Modul pasangan-aturan terdiri dari lima gen, dan semuanya, seperti yang Anda duga, saling menekan. Masing-masing gen ini bekerja baik di semua genap atau di semua segmen ganjil - protein yang dikodekan oleh mereka memecah lebih cepat daripada protein dari kelompok celah, dan strip yang sesuai lebih sempit. Mematikan sebagian gen dari kelompok ini memberikan embrio yang dipersingkat, di mana semua genap atau semua segmen ganjil menghilang.
Hubungan antara gen gap dan gen polaritas telur. Bcd - bicoid, hb - bongkok, cad - caudal, Kr - Kruppel, kni - knirps, gt - giant, tll - berekor.Penindasan gen masing-masing saja akan menandai embrio menjadi beberapa garis, tetapi lokasi pita aktivitas dari empat gen relatif satu sama lain dan ujung embrio akan sangat bervariasi. Agar strip selalu ada di tempat (misalnya, untuk knirps - satu strip di kepala, yang lain di bagian belakang antara kruppel dan strip raksasa), Anda memerlukan sinyal kontrol kedua - gradien bicoid dan protein polaritas telur lainnya. Fungsi input gen gap untuk regulator ini memiliki maksimum pada berbagai level sinyal input (kami memperoleh fungsi input seperti itu di akhir artikel kedua ). Oleh karena itu, gen-gen celah masing-masing dihidupkan oleh tingkat protein bicoid, hunchback, dan caudal mereka, dan zona aktivitas mereka secara alami terletak dari kepala ke ekor embrio.Setelah gen gap membuat tanda kasar embrio menjadi 7 pita (mereka tidak bertepatan dengan segmen atau kepala-dada-perut), modul berikutnya dihidupkan - aturan pasangan ("aturan aturan pasangan"). Itu sudah membagi embrio menjadi segmen, yang seharusnya 17. Modul pasangan-aturan terdiri dari lima gen, dan semuanya, seperti yang Anda duga, saling menekan. Masing-masing gen ini bekerja baik di semua genap atau di semua segmen ganjil - protein yang dikodekan oleh mereka memecah lebih cepat daripada protein dari kelompok celah, dan strip yang sesuai lebih sempit. Mematikan sebagian gen dari kelompok ini memberikan embrio yang dipersingkat, di mana semua genap atau semua segmen ganjil menghilang.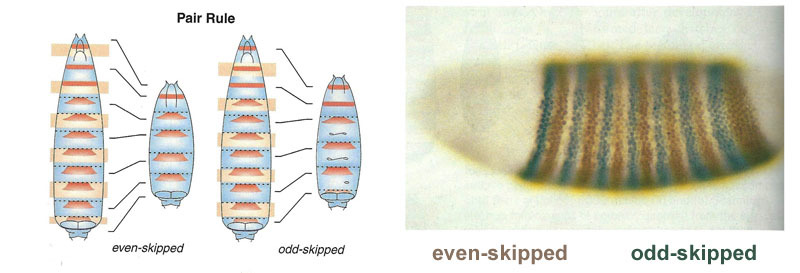 Zona aktivitas gen dari kelompok aturan pasangan. Foto di sebelah kanan memiliki 14 strip, bukan 17, karena tiga sistem penandaan lainnya berfungsi di tiga segmen pertama kepala.Modul aturan pasangan itu sendiri, tanpa partisipasi dari celah dan bicoid, dapat menandai embrio menjadi strip sempit, tetapi jumlah strip ini tidak stabil: segmen bisa 12, 18, dan bahkan 20. Biasanya, gen aturan pasangan sulit dikendalikan. kesenjangan gen, dan markup kasar pertama (untuk 7 zona) membantu secara akurat dan andal membuat markup akhir (untuk 14 + 3 segmen).Poin berikutnya dalam algoritma adalah untuk menandai bagian depan, tengah dan belakang dari setiap segmen. Ini dilakukan oleh gen polaritas segmen. Mereka juga saling menekan dan diaktifkan oleh modul jaringan gen yang lebih tinggi (pair-rule dan gap). Hanya parameter difusi dan pembusukan produk mereka yang dipilih sehingga strip sangat tipis - setengah segmen. Menekan mereka satu sama lain lebih lemah dan lebih terorganisir daripada dengan gen aturan pasangan. Oleh karena itu, strip aktivitas mereka sebagian tumpang tindih dan di setiap segmen sering "garis" strip berbeda tumpang tindih diperoleh.Mematikan gen polaritas segmen biasanya tidak menyebabkan pemendekan embrio dan kehilangan bagian-bagiannya (seperti tipikal celah dan aturan pasangan), tetapi untuk efek yang lebih halus. Biasanya, dalam kasus ini, setengah dari setiap segmen di mana gen ini seharusnya bekerja berubah menjadi gambar cermin dari setengah lainnya. Ini terlihat jelas, karena larva lalat memiliki barisan duri kecil yang diarahkan ke belakang - untuk kenyamanan merangkak. Mutan dalam segmen polaritas gen memiliki barisan duri diarahkan ke depan.Akhirnya, giliran gen yang paling terkenal dan pertama kali ditemukan dari jaringan ini - modul hox. Pekerjaan mereka memberikan segmen individualitas sehingga antena dan belalai tumbuh di segmen kepala, dan kaki dan sayap tumbuh di segmen dada. Gen hox, tidak seperti modul sebelumnya, tidak menekan aktivitas satu sama lain dan dapat bekerja bersama dalam segmen embrio yang sama. Sifat aneh gen hox yang belum dijelaskan: mereka terletak pada kromosom dalam kelompok yang ketat, dan dalam urutan yang sama di mana mereka bekerja di tubuh embrio.
Zona aktivitas gen dari kelompok aturan pasangan. Foto di sebelah kanan memiliki 14 strip, bukan 17, karena tiga sistem penandaan lainnya berfungsi di tiga segmen pertama kepala.Modul aturan pasangan itu sendiri, tanpa partisipasi dari celah dan bicoid, dapat menandai embrio menjadi strip sempit, tetapi jumlah strip ini tidak stabil: segmen bisa 12, 18, dan bahkan 20. Biasanya, gen aturan pasangan sulit dikendalikan. kesenjangan gen, dan markup kasar pertama (untuk 7 zona) membantu secara akurat dan andal membuat markup akhir (untuk 14 + 3 segmen).Poin berikutnya dalam algoritma adalah untuk menandai bagian depan, tengah dan belakang dari setiap segmen. Ini dilakukan oleh gen polaritas segmen. Mereka juga saling menekan dan diaktifkan oleh modul jaringan gen yang lebih tinggi (pair-rule dan gap). Hanya parameter difusi dan pembusukan produk mereka yang dipilih sehingga strip sangat tipis - setengah segmen. Menekan mereka satu sama lain lebih lemah dan lebih terorganisir daripada dengan gen aturan pasangan. Oleh karena itu, strip aktivitas mereka sebagian tumpang tindih dan di setiap segmen sering "garis" strip berbeda tumpang tindih diperoleh.Mematikan gen polaritas segmen biasanya tidak menyebabkan pemendekan embrio dan kehilangan bagian-bagiannya (seperti tipikal celah dan aturan pasangan), tetapi untuk efek yang lebih halus. Biasanya, dalam kasus ini, setengah dari setiap segmen di mana gen ini seharusnya bekerja berubah menjadi gambar cermin dari setengah lainnya. Ini terlihat jelas, karena larva lalat memiliki barisan duri kecil yang diarahkan ke belakang - untuk kenyamanan merangkak. Mutan dalam segmen polaritas gen memiliki barisan duri diarahkan ke depan.Akhirnya, giliran gen yang paling terkenal dan pertama kali ditemukan dari jaringan ini - modul hox. Pekerjaan mereka memberikan segmen individualitas sehingga antena dan belalai tumbuh di segmen kepala, dan kaki dan sayap tumbuh di segmen dada. Gen hox, tidak seperti modul sebelumnya, tidak menekan aktivitas satu sama lain dan dapat bekerja bersama dalam segmen embrio yang sama. Sifat aneh gen hox yang belum dijelaskan: mereka terletak pada kromosom dalam kelompok yang ketat, dan dalam urutan yang sama di mana mereka bekerja di tubuh embrio.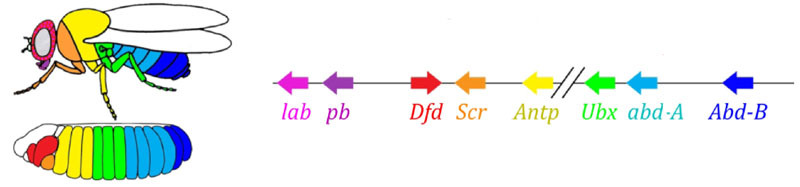 Penandaan embrio didasarkan pada prinsip “semakin jauh dari dada, semakin banyak gen hox dan semakin aktif itu”: aktivitas gen perut-A dimulai dengan 2 segmen perut, meningkat menjadi penuh dalam 4 dan berlanjut pada level maksimum di semua segmen berikutnya; Gen perut-B bekerja dari segmen ke-5 sampai ke-8 perut, meningkatkan aktivitas di setiap segmen berikutnya. Gen ultrabithorax termasuk dalam setengah kekuatan di segmen toraks ketiga dan pada kapasitas penuh di seluruh perut. Ketika dimatikan dalam mutan bithorax, sel-sel dari segmen 3 thoracic tidak tahu bahwa mereka adalah segmen 3 thoracic, mereka memiliki set gen hox yang sama seperti pada 2 thoracic (mis. Antennapedia saja) dan pengembangan dari 3 segmen thoracic dilanjutkan bersama program 2 dada - dengan sayap, bukan kumbang darat dan otot terbang yang kuat.Jika Anda mematikan semua gen hox di segmen terpisah, itu akan menjadi jelek, tetapi sebagian besar akan terlihat seperti 1 dada, dengan kaki dan tanpa sayap. Ini karena nenek moyang serangga yang jauh seperti trilobita atau kaki seribu. Mereka memiliki banyak segmen yang identik dengan kaki. Selanjutnya, tubuh mereka memendek, bagian belakang kehilangan kaki, menjadi perut, dan beberapa segmen depan bergabung dengan kepala dan kaki mereka menjadi bagian dari alat oral (mandibula, rahang atas dan bibir bawah).
Penandaan embrio didasarkan pada prinsip “semakin jauh dari dada, semakin banyak gen hox dan semakin aktif itu”: aktivitas gen perut-A dimulai dengan 2 segmen perut, meningkat menjadi penuh dalam 4 dan berlanjut pada level maksimum di semua segmen berikutnya; Gen perut-B bekerja dari segmen ke-5 sampai ke-8 perut, meningkatkan aktivitas di setiap segmen berikutnya. Gen ultrabithorax termasuk dalam setengah kekuatan di segmen toraks ketiga dan pada kapasitas penuh di seluruh perut. Ketika dimatikan dalam mutan bithorax, sel-sel dari segmen 3 thoracic tidak tahu bahwa mereka adalah segmen 3 thoracic, mereka memiliki set gen hox yang sama seperti pada 2 thoracic (mis. Antennapedia saja) dan pengembangan dari 3 segmen thoracic dilanjutkan bersama program 2 dada - dengan sayap, bukan kumbang darat dan otot terbang yang kuat.Jika Anda mematikan semua gen hox di segmen terpisah, itu akan menjadi jelek, tetapi sebagian besar akan terlihat seperti 1 dada, dengan kaki dan tanpa sayap. Ini karena nenek moyang serangga yang jauh seperti trilobita atau kaki seribu. Mereka memiliki banyak segmen yang identik dengan kaki. Selanjutnya, tubuh mereka memendek, bagian belakang kehilangan kaki, menjadi perut, dan beberapa segmen depan bergabung dengan kepala dan kaki mereka menjadi bagian dari alat oral (mandibula, rahang atas dan bibir bawah).Model matematika dari jaringan genetik untuk menandai embrio
Dalam kasus operon laktosa dan loop tautan langsung pada E. coli, dimungkinkan untuk membuat model matematika yang cukup sederhana dengan sejumlah parameter moderat yang memprediksi perilaku gen untuk setiap sinyal input. Untuk operon laktosa, itu sudah cukup:- empat konstanta pengikat (represor dan aktivator dengan DNA, laktosa dengan represor, cAMP dengan aktivator)- empat parameter stoikiometri (4 laktosa per 1 represor, 2 aktivator cAMP 1, 1 represor dan aktivator per operon)- dua parameter pengaturan (pengaruh represor dan aktivator pada kemungkinan pendaratan RNA polimerase)- tingkat dasar aktivitas operon (ketika tidak ada represor atau aktivator)- penundaan transkripsiTotal 12 parameter.Sekarang mari kita lihat wilayah regulasi, katakanlah, gen Hox. Dalam diagram di bawah ini, salah satu blok pengatur gen ab-B. Bilah berwarna menunjukkan situs pengikatan protein regulatori Seluruh blok ini memiliki ukuran sekitar 1700 nukleotida - 10 kali lebih lama dari wilayah regulasi operasi laktosa. Ia memiliki 39 lokasi penanaman untuk 7 protein regulator berbeda dari semua blok superior jaringan gen ini. Dan abdominal-B memiliki empat blok regulasi seperti itu. Mereka menyala secara berurutan dari segmen perut ke-5 dan ke-8 dan menciptakan peningkatan bertahap dalam kadar protein B-perut di segmen-segmen ini. Yaitu, untuk model mat gen ini saja, dengan cara yang baik, Anda membutuhkan lebih dari 100 konstanta pengikat, jumlah parameter regulasi yang sama, dan jumlah parameter yang tidak diketahui untuk menggambarkan interaksi antara input yang berbeda - dalam kasus terburuk, hingga 1.500 interaksi berpasangan antara semua situs pendaratan protein dalam satu unit pengatur.Yaitu, model lengkap dari jaringan gen ini tentunya akan memiliki lebih dari seribu parameter. Dan ini adalah masalah utama: tidak ada begitu banyak sinyal keluaran yang dapat kita ukur dalam percobaan - pada kenyataannya, hanya tingkat masing-masing dari 25 protein dan distribusinya sepanjang embrio. Oleh karena itu, tidak ada yang mencoba mensimulasikan jaringan ini dengan granularity yang sama dengan operon laktosa.Anda dapat mencoba menyederhanakan tugas dan menjelaskan efek dari masing-masing protein pengatur pada gen dengan hanya tiga parameter (pengikatan, stoikiometri, dan parameter pengaturan). Anda dapat memilih parameter ini (misalnya, menggunakan rantai Markov) sehingga model secara akurat mereproduksi strip nyata aktivitas gen. Untuk dua blok pertama jaringan (gen polaritas telur dan gen gap) model seperti itu bahkan dibuat ( misalnya ).Masalahnya adalah bahwa permulaan yang berbeda dari algoritma pemilihan memberikan parameter model yang berbeda - tetapi mereka semua bekerja dengan benar dan mereproduksi aktivitas gen dalam tunas lalat nyata! Model-model ini memiliki fitur umum (misalnya, semua gen celah saling menekan dan diaktifkan oleh protein induk), tetapi tidak ada perbedaan yang kurang dari kesamaan.Dan karena berbagai upaya pemodelan memberikan hasil yang berbeda, tidak mungkin bahwa setidaknya satu di antaranya bertepatan dengan kenyataan. Lebih buruk lagi, model seperti itu mereproduksi operasi normal, tetapi kehilangan banyak dalam memprediksi hasil mutasi.Pada bagian selanjutnya, kita akan berbicara tentang mengapa masalah ini dengan studi jaringan gen tidak dapat dihindari dalam biologi (dan jaringan penandaan embrio Drosophila adalah salah satu yang paling sederhana untuk dipelajari!), Apa yang dapat dilakukan tentang hal ini, dan insinyur apa yang harus dipelajari dari ahli biologi.
Bilah berwarna menunjukkan situs pengikatan protein regulatori Seluruh blok ini memiliki ukuran sekitar 1700 nukleotida - 10 kali lebih lama dari wilayah regulasi operasi laktosa. Ia memiliki 39 lokasi penanaman untuk 7 protein regulator berbeda dari semua blok superior jaringan gen ini. Dan abdominal-B memiliki empat blok regulasi seperti itu. Mereka menyala secara berurutan dari segmen perut ke-5 dan ke-8 dan menciptakan peningkatan bertahap dalam kadar protein B-perut di segmen-segmen ini. Yaitu, untuk model mat gen ini saja, dengan cara yang baik, Anda membutuhkan lebih dari 100 konstanta pengikat, jumlah parameter regulasi yang sama, dan jumlah parameter yang tidak diketahui untuk menggambarkan interaksi antara input yang berbeda - dalam kasus terburuk, hingga 1.500 interaksi berpasangan antara semua situs pendaratan protein dalam satu unit pengatur.Yaitu, model lengkap dari jaringan gen ini tentunya akan memiliki lebih dari seribu parameter. Dan ini adalah masalah utama: tidak ada begitu banyak sinyal keluaran yang dapat kita ukur dalam percobaan - pada kenyataannya, hanya tingkat masing-masing dari 25 protein dan distribusinya sepanjang embrio. Oleh karena itu, tidak ada yang mencoba mensimulasikan jaringan ini dengan granularity yang sama dengan operon laktosa.Anda dapat mencoba menyederhanakan tugas dan menjelaskan efek dari masing-masing protein pengatur pada gen dengan hanya tiga parameter (pengikatan, stoikiometri, dan parameter pengaturan). Anda dapat memilih parameter ini (misalnya, menggunakan rantai Markov) sehingga model secara akurat mereproduksi strip nyata aktivitas gen. Untuk dua blok pertama jaringan (gen polaritas telur dan gen gap) model seperti itu bahkan dibuat ( misalnya ).Masalahnya adalah bahwa permulaan yang berbeda dari algoritma pemilihan memberikan parameter model yang berbeda - tetapi mereka semua bekerja dengan benar dan mereproduksi aktivitas gen dalam tunas lalat nyata! Model-model ini memiliki fitur umum (misalnya, semua gen celah saling menekan dan diaktifkan oleh protein induk), tetapi tidak ada perbedaan yang kurang dari kesamaan.Dan karena berbagai upaya pemodelan memberikan hasil yang berbeda, tidak mungkin bahwa setidaknya satu di antaranya bertepatan dengan kenyataan. Lebih buruk lagi, model seperti itu mereproduksi operasi normal, tetapi kehilangan banyak dalam memprediksi hasil mutasi.Pada bagian selanjutnya, kita akan berbicara tentang mengapa masalah ini dengan studi jaringan gen tidak dapat dihindari dalam biologi (dan jaringan penandaan embrio Drosophila adalah salah satu yang paling sederhana untuk dipelajari!), Apa yang dapat dilakukan tentang hal ini, dan insinyur apa yang harus dipelajari dari ahli biologi.