
Terlepas dari adanya hambatan dan kesulitan yang jelas yang terkadang menghalangi pengembangan dan implementasi produk rekayasa genetika, abad ke-21 tidak dapat dibayangkan tanpa hasil dari teknologi yang penting dan beragam ini dalam gudang seorang ahli biologi modern. Organisme yang paling umum digunakan dalam GI adalah bakteri.
Apa itu GI dan mengapa kita membutuhkannya? Mengapa bakteri begitu populer dengan para insinyur genetika? Dalam bentuk apa cara termudah untuk memasukkan gen yang diinginkan ke dalam bakteri? Kesulitan apa yang bisa ditemui ketika bekerja dengan organisme ini? Apa yang terjadi sebelumnya: penciptaan bakteri rekayasa genetika pertama atau penemuan struktur DNA dan genom? Baca tentang ini dan banyak lagi di bawah kucing.
0. Program pendidikan singkat dalam biologi
Paragraf ini memberikan deskripsi singkat tentang apa yang disebut
Dogma Pusat biologi molekuler . Jika Anda memiliki pengetahuan dasar dalam biologi molekuler, silakan lompat ke langkah 1.
 Dogma sentral biologi molekuler dalam satu gambar
Dogma sentral biologi molekuler dalam satu gambarJadi mari kita mulai. Semua informasi tentang semua tahap perkembangan dan sifat-sifat organisme apa pun, baik
prokariota (bakteri),
archaea atau
eukariota (semua sisanya adalah tunggal dan multiseluler), dikodekan dalam DNA genom, yang merupakan kompleks dari dua rantai polinukleotida yang saling melengkapi satu sama lain, membentuk heliks ganda ( nukleotida DNA komplementer: AT dan GC). Kromosom eukariotik adalah molekul DNA beruntai ganda linier, dan kromosom prokariotik diulang. Seringkali, gen membentuk hanya sebagian kecil dari seluruh genom (pada manusia - sekitar 1,5%).
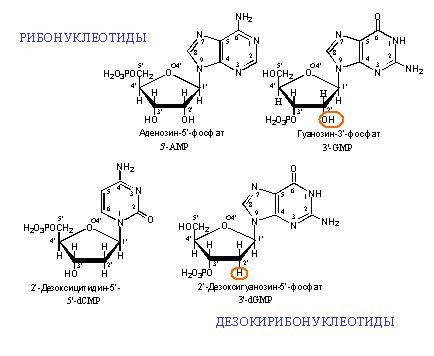 Contoh monomer DNA dan RNA. "Deoksi" dalam nama DNA berarti tidak adanya atom oksigen pada posisi 2 '(pada gambar, posisi 2' dilingkari merah).
Contoh monomer DNA dan RNA. "Deoksi" dalam nama DNA berarti tidak adanya atom oksigen pada posisi 2 '(pada gambar, posisi 2' dilingkari merah).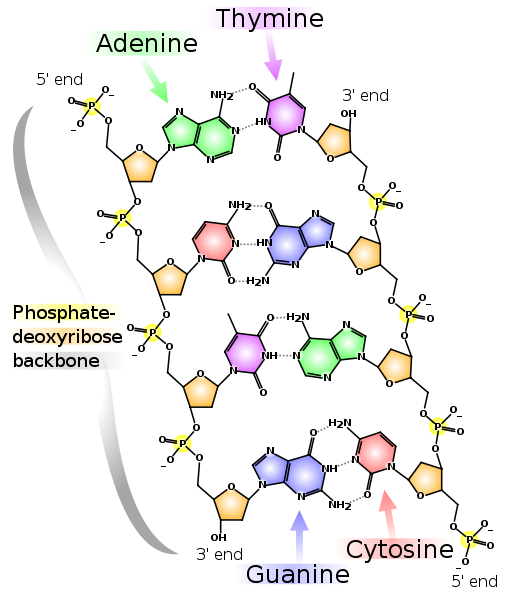 Dua untai DNA komplementer. Garis putus-putus menunjukkan ikatan hidrogen antara basis. Seperti dapat dilihat, adenin dan timin membentuk dua ikatan hidrogen antara satu sama lain, dan guanin dan sitosin membentuk tiga. Oleh karena itu, ikatan GC lebih kuat dan bagian DNA untaian ganda kaya GC lebih sulit untuk dipisahkan menjadi dua rantai.
Dua untai DNA komplementer. Garis putus-putus menunjukkan ikatan hidrogen antara basis. Seperti dapat dilihat, adenin dan timin membentuk dua ikatan hidrogen antara satu sama lain, dan guanin dan sitosin membentuk tiga. Oleh karena itu, ikatan GC lebih kuat dan bagian DNA untaian ganda kaya GC lebih sulit untuk dipisahkan menjadi dua rantai.Perhatikan bahwa setiap rantai memiliki 5'-end dan 3'-end. Dapat dilihat bahwa di dekat ujung 5'-rantai kiri adalah 3'-ujung kanan dan sebaliknya, oleh karena itu rantai disebut "antiparalel." RNA juga memiliki ujung 5'- dan 3'-akhir. Posisi 5 'dan 3' sendiri dipilih untuk menunjukkan awal dan akhir karena melalui mereka ikatan kovalen terbentuk dalam rantai DNA dan RNA.
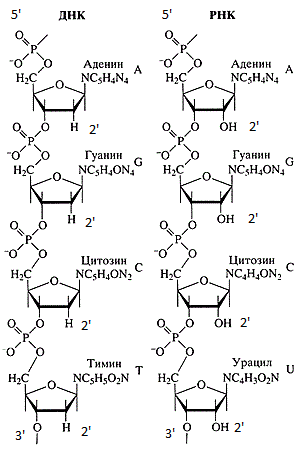 Rantai DNA dan RNA.
Rantai DNA dan RNA.Sekuens DNA dan RNA selalu direkam dari ujung 5'ke ujung 3'. Ada sejumlah alasan untuk ini:
- Sintesis rantai DNA dan RNA baru dimulai dari ujung 5'-end ( DNA polimerase (enzim yang mensintesis rantai DNA komplementer pada matriks DNA atau RNA) dan RNA polimerase (enzim yang mensintesis rantai RNA komplementer pada DNA atau matriks RNA) arah 3 '-> 5', sehingga rantai baru disintesis dalam arah 5 '-> 3');
- Ribosom membaca kodon, bergerak di sepanjang mRNA ke arah 5 '-> 3';
- Urutan asam amino ditulis dalam rantai pengkodean DNA dalam arah 5 '-> 3' (bagian penting dari mRNA adalah salinan yang tepat dari daerah pengkodean DNA dengan timin diganti oleh urasil dan dengan kelompok hidroksil (-OH) alih-alih hidrogen pada posisi 2 ', tentu saja);
- Akhirnya, hanya nyaman untuk memiliki aturan perekaman yang diterima secara umum.
Gen adalah bagian dari DNA genom yang menentukan urutan nukleotida dari molekul RNA:
- Encoding RNA: messenger RNA (mRNA), di mana urutan asam amino dari protein yang sesuai dikodekan sebagai kodon. Anda juga dapat menemukan nama "RNA informasi", maka singkatannya tampak seperti "mRNA";
- Non-coding RNA: transport RNA, RNA ribosom dan lainnya.
Peran tRNA adalah untuk mengirimkan asam amino ke kompleks mRNA-ribosom. Selain itu, tRNA yang bertanggung jawab untuk pengenalan kodon mRNA, untuk ini, masing-masing tRNA mencakup apa yang disebut "antikodon" - triplet yang saling melengkapi dengan kodon mRNA.
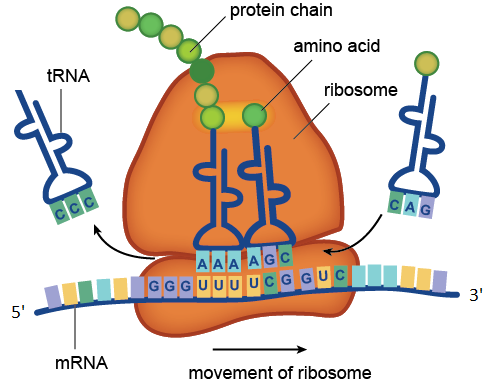 Proses terjemahan dikatalisasi oleh ribosom. Dalam gambar, kodon UUU dan UCG yang terkandung dalam mRNA diakui oleh antikodon AAA dan AGC yang terkandung dalam molekul tRNA. Transport RNA dengan anticodon CCC telah memberikan asam amino untuk rantai protein yang sedang tumbuh, dan tRNA dengan anticodon CAG menunggu dalam antrian. Plot molekul mRNA yang ditunjukkan pada gambar terdiri dari empat kodon: GGGUUUCUCGGUC. Kodon GGG sesuai dengan glisin asam amino, UUU menjadi fenilalanin, UCG untuk serin, GUC ke valin. Jadi wilayah mRNA ini mengkode fragmen protein dengan sekuens asam amino glikin-fenilalanin-serin-valin.
Proses terjemahan dikatalisasi oleh ribosom. Dalam gambar, kodon UUU dan UCG yang terkandung dalam mRNA diakui oleh antikodon AAA dan AGC yang terkandung dalam molekul tRNA. Transport RNA dengan anticodon CCC telah memberikan asam amino untuk rantai protein yang sedang tumbuh, dan tRNA dengan anticodon CAG menunggu dalam antrian. Plot molekul mRNA yang ditunjukkan pada gambar terdiri dari empat kodon: GGGUUUCUCGGUC. Kodon GGG sesuai dengan glisin asam amino, UUU menjadi fenilalanin, UCG untuk serin, GUC ke valin. Jadi wilayah mRNA ini mengkode fragmen protein dengan sekuens asam amino glikin-fenilalanin-serin-valin.RNA ribosom adalah komponen yang sangat diperlukan dari ribosom. Fungsi utama rRNA adalah untuk memastikan proses penerjemahan: terlibat dalam membaca informasi dari mRNA menggunakan molekul adaptor tRNA dan mengkatalisis pembentukan ikatan peptida antara asam amino yang melekat pada tRNA dan rantai protein yang sedang tumbuh.
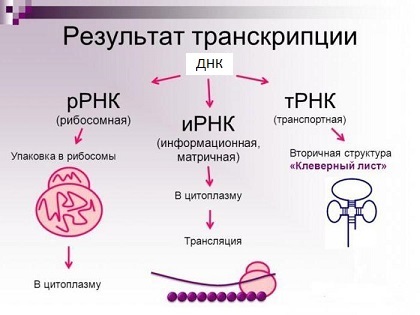 Jenis utama molekul RNA (pada kenyataannya, ada lebih banyak lagi).
Jenis utama molekul RNA (pada kenyataannya, ada lebih banyak lagi).Sebuah protein, di sisi lain, adalah rantai asam amino yang secara kovalen dihubungkan bersama melalui ikatan peptida (Anda dapat melihat apa yang ada di spoiler sedikit lebih jauh). Setelah sintesis, rantai asam amino harus mengambil struktur spasial tertentu - "
konformasi " (
mereka sudah memberi tahu saya tentang struktur spasial protein
di Geektimes ). Selain itu, banyak protein besar sebenarnya terdiri dari beberapa protein yang dikombinasikan oleh interaksi hidrofobik dan ikatan hidrogen menjadi struktur tunggal yang stabil. Dalam hal ini, masing-masing "protein pembangun" disebut "subunit", dan protein besar yang dihasilkan disebut "multisubunit".
20 asam amino yang membentuk protein  Kompleks ribosom. Gambar itu diambil dari publikasi OlegKovalevskiy "pencetakan 3D model molekul protein" .
Kompleks ribosom. Gambar itu diambil dari publikasi OlegKovalevskiy "pencetakan 3D model molekul protein" .Dalam kasus gen yang mengkode protein, proses penguraian informasi genetik terlihat seperti ini:
- RNA polimerase mengenali promotor dan mengikatnya (jika "terbuka", kami akan membahas lebih lanjut pengaturan aktivitas promotor);
- Berdasarkan prinsip saling melengkapi, enzim RNA polimerase pada matriks DNA mensintesis "blank" RNA templat (pre-mRNA, dalam eukariota) atau mRNA fungsional siap pakai (dalam prokariota). Proses ini disebut "transkripsi" ;
- (hanya dalam eukariota) Molekul pra-mRNA mengalami modifikasi ("matang") dan menjadi mRNA fungsional;
- mRNA dikenali oleh ribosom , enzim yang menerjemahkan kode triplet mRNA dan, berdasarkan itu, mensintesis peptida / protein. Asam amino dari mana ribosom membangun protein dikirim dalam kompleks dengan transport RNA ( tRNA ). Proses ini disebut "penyiaran" ;
- Peptida / protein dapat mengalami modifikasi pasca-translasi ("pematangan" dengan analogi dengan mRNA) dan menjadi fungsional. Faktor penting adalah bahwa sistem modifikasi eukariota pasca-translasi jauh lebih kompleks dan beragam daripada prokariota, oleh karena itu, tidak setiap protein eukariotik dapat disintesis dengan benar oleh bakteri.
Selain daerah pengkodean, genom berisi banyak fragmen yang juga berpartisipasi dalam transkripsi dengan satu atau lain cara. Plot yang terletak dekat dengan gen dan disebut promotor diakui oleh RNA polimerase (mereka mengatakan bahwa gen berada di bawah kendali promotor ini). Promotor yang berbeda diakui oleh RNA polimerase yang berbeda. Misalnya, gen di bawah kendali promotor
bakteriofag tidak akan ditranskripsi menjadi bakteri jika RNA polimerase bakteriofag yang sesuai tidak disintesis di dalamnya.
Umumnya .
Setiap gen juga dapat memiliki beberapa sekuens pengatur, yang dapat ditempatkan baik secara langsung di dekat promotor (atau bahkan tumpang tindih dengannya), atau pada jarak puluhan ribu pasangan nukleotida darinya. Elemen
penambah transkripsi disebut
"peningkat," elemen penekan transkripsi disebut peredam, dan protein yang berinteraksi dengannya disebut
faktor transkripsi . Meskipun juga biasa menyebut faktor transkripsi komponen yang diperlukan dari kompleks inisiasi transkripsi, yang tanpanya transkripsi tidak mungkin secara prinsip. Faktanya adalah bahwa hanya untuk memulai sintesis molekul RNA pada matriks DNA dalam eukariota dan archaea, perakitan seluruh kompleks supramolekul diperlukan. Kompleks yang paling sederhana termasuk holoenzyme RNA polimerase dan enam yang disebut
"faktor transkripsi umum" (TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIF, dan TFIIH). Kompleks itu sendiri disebut
"Kompleks preinisiasi transkripsi" (
video , setiap komponen kompleks disorot dalam satu warna atau lainnya).
Kompleks transkripsi prokariotik benar-benar berbeda, sehingga tidak ada gunanya menanamkan gen eukariotik dengan promotor eukariotik dalam bakteri. Analog prokariotik dari faktor transkripsi umum eukariota dan archaea dapat disebut protein yang disebut
"faktor sigma" .
 Kompleks transkripsi prokariotik. Huruf yang ditunjukkan pada gambar adalah sebutan umum yang diterima dari subunit yang sesuai. σ70 - faktor sigma gen rumah tangga E. coli
Kompleks transkripsi prokariotik. Huruf yang ditunjukkan pada gambar adalah sebutan umum yang diterima dari subunit yang sesuai. σ70 - faktor sigma gen rumah tangga E. coliGenom prokariota dan eukariota memiliki banyak kesamaan, dan Dogma Tengah dari Biologi Molekuler, yang disebutkan sebelumnya, berlaku untuk kedua kerajaan. Namun, ada juga banyak perbedaan signifikan. Sebagai contoh, bakteri dicirikan oleh sistem operon - gen yang dikelompokkan bersama yang berpartisipasi dalam proses yang sama dan tidak ditranskripsi secara terpisah, tetapi sebagai bagian dari satu mRNA yang panjang. Dalam eukariota, semuanya benar-benar berbeda: gen yang terlibat dalam satu proses tersebar di kromosom yang berbeda, dan gen itu sendiri dibagi menjadi fragmen pengkodean
ekson oleh daerah
intron yang tidak mengkode. Dalam hal ini, pada awalnya gen ditranskrip sepenuhnya, dan kemudian, sudah pada tahap RNA, intron dieksisi, dan ekson cross-link untuk membentuk mRNA pengkodean. Proses ini disebut
splicing . Pada saat yang sama, tidak semua ekson yang tersedia dapat dijahit ke mRNA yang sudah jadi, tetapi hanya sebagian dari mereka, dalam hal ini, berbicara tentang
“penyambungan alternatif” . Dengan demikian, sel eukariotik dapat mensintesis beberapa protein, sambil menyalin gen yang sama. Di antara hal-hal lain, ini memberikan konsekuensi yang sangat penting: sering kali tidak masuk akal untuk memasukkan gen eukariotik ke dalam bakteri "seperti yang ada di dalam kromosom", karena bakteri tidak dapat melakukan splicing.
Ada perbedaan penting lainnya. Prokariota dicirikan oleh adanya bahan genetik berbasis DNA di luar cincin "kromosom", yang disebut
"plasmid" - molekul DNA sirkuler ganda-untai kecil. Selain itu, prokariota kekurangan organel, termasuk nukleus: semua komponen sel bakteri bebas untuk bepergian ke seluruh ruang intraseluler. Eukariota, bagaimanapun, tidak memiliki plasmid, tetapi ada
plastid dan
mitokondria dalam genom yang termasuk plasmid (menurut hipotesis yang paling kuat, plastid dan mitokondria adalah "keturunan" dari arsitektur prokariotik dari genom cyanobacteria dan bakteri yang terperangkap di dalamnya oleh proto-unicellular kuno). Selain itu, keberadaan nukleus dan kompartemen intraseluler lainnya dikelilingi oleh membrannya sendiri sudah khas untuk eukariota. Oleh karena itu, rekayasa genetika sel eukariotik memerlukan pendekatan yang berbeda dari rekayasa genetika bakteri.
Kode genetik itu sendiri disusun sebagai berikut. Setiap gen / ekson terdiri dari satu set triplet / kodon - urutan tiga nukleotida di antaranya tidak ada celah. Organisasi triplet berlaku untuk gen dalam DNA dan untuk bagian pengkodean mRNA. Dalam proses penerjemahan, transport RNAs (tRNAs) yang membawa asam amino spesifik “mengenali” kembar tiga tiga huruf yang sesuai. Ribosom memutus asam amino dari tRNA dan menempelkannya ke rantai asam amino yang sedang tumbuh, yang pada akhir terjemahan akan segera menjadi protein matang, berfungsi penuh, atau sebelum itu juga mengalami serangkaian modifikasi. Dalam hal ini, hanya satu asam amino yang berhubungan dengan setiap triplet, tetapi beberapa kodon yang berbeda dapat berhubungan dengan satu asam amino. Ini bisa dimengerti, karena dalam kode genetik standar ada 61 kodon pengkode, dan
hanya ada 20 asam amino proteinogenik (kodon total, tentu saja, 4 * 4 * 4 = 64, tetapi tiga di antaranya bukan pengkodean, sebaliknya mereka berfungsi sebagai sinyal untuk menghentikan terjemahan dan disebut " hentikan kodon ”).
 Kodon dalam kode genetik standar. Berkat Wikipedia untuk fotonya.
Kodon dalam kode genetik standar. Berkat Wikipedia untuk fotonya.Jadi, protein adalah elemen-elemen yang merupakan penghubung terakhir dalam rantai antara DNA genom dan sifat-sifat tubuh, yang disebut
"fenotip" . Oleh karena itu, untuk mengubah karakteristik organisme yang penting bagi kita, kita perlu mengubah DNA-nya sedemikian rupa sehingga, sebagai akibatnya, protein tertentu muncul dalam selnya, yang akan memberi kita hasil target. Ini adalah ide dasar dari semua rekayasa genetika.
1) Untuk tujuan apa bakteri digunakan dalam rekayasa genetika dan mengapa demikian
Jadi, kami menemukan bagaimana dan mengapa urutan genomik DNA memengaruhi sifat dan karakteristik tubuh. Tentu saja, akan sangat baik jika sifat tersebut sepenuhnya ditentukan oleh satu gen saja - memasukkan sebuah fragmen kecil bukan lagi masalah serius. Misalnya, sering kali resistensi tanaman terhadap herbisida atau hama ditentukan oleh satu gen, sehingga membuat varietas dengan resistensi yang diinginkan dalam kasus-kasus seperti itu tidak sulit (sebagai lawan untuk membawa tanaman seperti itu ke pasar). Hal yang sama berlaku untuk banyak resistensi antibiotik terhadap bakteri (pada kenyataannya, bakteri memiliki banyak mekanisme perlindungan terhadap antibiotik, tetapi mereka bekerja secara independen). Contoh sebaliknya adalah, misalnya, upaya oleh para ilmuwan untuk mengajarkan tanaman menyerap nitrogen dari atmosfer. Faktanya adalah bahwa satu-satunya sumber nitrogen untuk tanaman adalah tanah di mana senyawa yang mengandung nitrogen yang cocok untuk asimilasi oleh tanaman disintesis oleh mikroorganisme (atau diperkenalkan dalam bentuk pupuk oleh tukang kebun yang peduli atau anjing yang lewat). Jelas, menciptakan tanaman dengan mekanisme nutrisi alternatif akan sangat bermanfaat bagi pertanian. Namun, sayangnya, proses ini begitu rumit sehingga masalah "transfer" dari mikroorganisme ke pabrik belum terpecahkan sejauh ini.
Akhirnya, jika tujuan kami adalah untuk mendapatkan protein untuk tujuan tertentu (untuk mempelajari struktur dan fungsi protein, untuk membuat persiapan medis atau reagen laboratorium berdasarkan itu, dll), maka, jelas, kami juga cukup senang dengan integrasi satu gen tunggal ke dalam sel, yang dalam hal ini, biasanya disebut "organisme produsen".
Bakteri dalam rekayasa genetika adalah bahan sumber potensial untuk membuat:
- penghasil protein yang kita butuhkan pada skala laboratorium atau industri;
- agen aktif dalam transformasi kimia tertentu dari satu senyawa menjadi senyawa lain, apakah itu merupakan proses fermentasi dalam industri makanan, penciptaan kondisi yang lebih menguntungkan bagi pertumbuhan tanaman melalui pengenalan "produsen pupuk bakteri" di tanah atau pembuangan skrap baja;
- klonotek gen (sebuah topik, deskripsi yang bagus akan meningkatkan ukuran artikel menjadi tidak senonoh);
- obat yang signifikan secara medis, misalnya, untuk mengembalikan mikroflora pada saluran pencernaan;
- strain bakteri Agrobacterium tumefaciens untuk modifikasi genetik tanaman selanjutnya.
* Saya bisa melupakan sesuatu, jadi tambahan di komentar dipersilahkan.
Fakta menarik adalah bahwa percobaan sukses pertama di bidang rekayasa genetika bakteri terjadi jauh sebelum karya tengara Watson dan Crick. Selain itu, berdasarkan percobaan ini, fakta bahwa informasi tersebut terkandung dalam DNA terbukti, setelah itu para ilmuwan tidak dapat menghabiskan waktu mereka pada hipotesis tentang RNA dan protein.
Pekerjaan ini, yang dilakukan pada tahun 1944, dikenal sebagai
Eksperimen Avery, MacLeod, dan McCarthy , berdasarkan
karya Frederick Griffith , di mana selama itu ditemukan bahwa infeksi strain pneumokokus patogen dan non-patogen yang hidup menyebabkan perkembangan penyakit, sementara secara individual mereka tidak menyebabkan gejala yang signifikan. Dari percobaan ini, disimpulkan bahwa bakteri yang terbunuh mampu mentransmisikan sesuatu kepada “kolega” non-patogen, dan karenanya menjadi berbahaya. Tapi apa yang mereka saling berikan? Pada 1944, ada tiga kandidat utama: DNA, RNA, dan protein. Untuk membangun pembawa, percobaan elegan dilakukan: pada saat itu enzim yang mampu menghancurkan DNA secara terpisah (DNase), secara terpisah RNA (RNase) dan secara terpisah protein (proteinase) sudah tersedia. Terlihat bahwa transfer sifat-sifat patogen tidak hanya terjadi pada kasus-kasus tersebut ketika persiapan strain patogen yang mati diobati dengan DNase dan tidak tergantung pada perawatan obat dengan RNase dan proteinase.
Dengan demikian, terbukti bahwa DNA adalah pembawa informasi tentang tanda-tanda tersebut. Selain itu, jelas ditunjukkan bahwa penetrasi spontan molekul DNA asing ke dalam sel bakteri dimungkinkan.
Mengapa bakteri begitu populer dengan kekurangan yang jelas (misalnya, kurangnya modifikasi pasca-translasi eukariotik)? Semuanya sederhana.
Mereka bersahaja dalam operasi, mudah digunakan dan tidak memerlukan media nutrisi yang mahal.2) Bagaimana suatu konstruksi genetik dibuat yang dimasukkan ke dalam bakteri
Rekayasa genetika modern bakteri terutama adalah pengenalan vektor plasmid (plasmid bakteri termodifikasi yang mengandung gen target dan serangkaian elemen lain yang diperlukan, yang akan dibahas di bawah). Mengubah kromosom bakteri kurang khas, tetapi prosedur ini juga tidak aneh: misalnya, gen T7 bacteriophage RNA polimerase dimasukkan ke dalam kromosom E. coli menggunakan vektor berdasarkan pada profage λ selama pembuatan salah satu strain yang populer di plak laboratorium. Ada tiga alasan mengapa seorang peneliti akan sering memilih untuk memperkenalkan gen dalam vektor plasmid:Vektor plasmid tipikal untuk bekerja dengan bakteri adalah molekul DNA sirkuler ganda untai ganda yang membawa gen protein target di bawah kendali promotor spesifik dan sejumlah gen dan elemen pengatur yang diperlukan, yang keberadaannya memastikan jumlah plasmid yang konstan dalam sel (“kontrol salin”). Jelas, bahkan dalam kasus sintesis mRNA yang sangat efisien, vektor tidak banyak berguna jika ada dalam bakteri dalam jumlah sepasang: dalam proses pembelahan, kemungkinan pembentukan sel anak tanpa plasmid yang diperlukan akan dangkal.Selain gen dan promotor, elemen utama vektor plasmid adalah:- ori adalah daerah asal replikasi plasmid. Diperlukan untuk mempertahankan jumlah konstan plasmid dan pewarisannya oleh sel anak;
- — , , , . , , (« »). , , . , .
, β- (GUS). , . , . — (GFP) ( GUS GFP );
- , ( — , — );
- — , ( ). , «» .
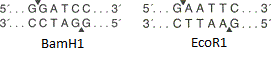 Angka menunjukkan situs pembatasan endonuklease BamH1 dan yang EcoR1 . Kedua enzim mengenali situs spesifik dari enam pasangan basa dan memperkenalkan istirahat beruntai tunggal di tempat yang berbeda (ditunjukkan oleh panah segitiga). Dalam hal ini, titik-titik putus rantai tidak bertepatan, yang berarti bahwa "ujung lengket" terbentuk (jika bertepatan, "ujung tumpul" terbentuk).
Angka menunjukkan situs pembatasan endonuklease BamH1 dan yang EcoR1 . Kedua enzim mengenali situs spesifik dari enam pasangan basa dan memperkenalkan istirahat beruntai tunggal di tempat yang berbeda (ditunjukkan oleh panah segitiga). Dalam hal ini, titik-titik putus rantai tidak bertepatan, yang berarti bahwa "ujung lengket" terbentuk (jika bertepatan, "ujung tumpul" terbentuk).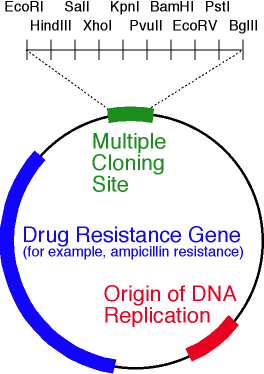 Skema vektor plasmid yang disederhanakan. Gambar tersebut menunjukkan ori, gen resistensi antibiotik dan polylinker yang mengandung 10 situs endonuklease restriksi.Nah, vektor ada di tangan kita. Bagaimana cara menanamkan gen di dalamnya? Lagi pula, di mana saya bisa mendapatkan gen ini?Misalkan kita tahu urutan nukleotida dari gen yang kita butuhkan. Kemudian lanjutkan sebagai berikut:
Skema vektor plasmid yang disederhanakan. Gambar tersebut menunjukkan ori, gen resistensi antibiotik dan polylinker yang mengandung 10 situs endonuklease restriksi.Nah, vektor ada di tangan kita. Bagaimana cara menanamkan gen di dalamnya? Lagi pula, di mana saya bisa mendapatkan gen ini?Misalkan kita tahu urutan nukleotida dari gen yang kita butuhkan. Kemudian lanjutkan sebagai berikut:- , ;
- .
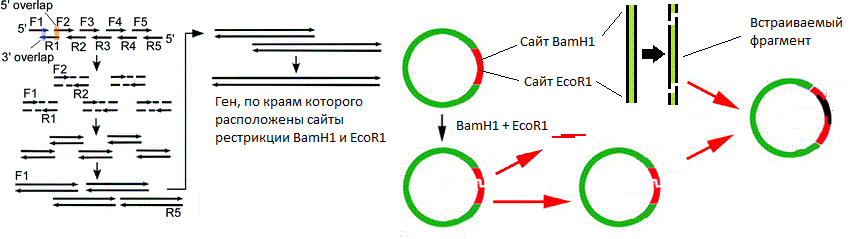 ( ). .Primer yang tumpang tindih disintesis sedemikian rupa sehingga setelah perakitan, gen berukuran penuh terbentuk, pada sisi-sisi di mana terdapat situs pemotongan dengan endonukleas yang sama, lokasi yang terletak di polylinker vektor. Dengan demikian, jika proses endonuklease yang diperlukan (dalam contoh saya adalah enzim-enzim BamH1 dan yang EcoR1 ) vektor dan gen yang dirakit, maka mereka membentuk ujung lengket yang dapat "mengenal satu sama lain" karena interaksi nukleotida komplementer ujung lengket. Tetapi pengenalan saja tidak cukup, karena sejauh ini ujung lengket saling menempel hanya karena ikatan hidrogen yang rapuh. Masalah ini diselesaikan dengan menambahkan campuran reaksi secara bersamaan memotong vektor, memotong gen dan enzim DNA ligase ., yang menghilangkan kerusakan rantai pada molekul DNA untai ganda.Faktor penting lain dalam perakitan gen adalah kenyataan bahwa frekuensi satu atau lain kodon berbeda untuk organisme yang berbeda, sementara di dalam sel biasanya ada lebih banyak tRNA yang sesuai dengan kodon yang lebih "populer". Karena banyak asam amino dikodekan oleh beberapa kodon, sangat mungkin bahwa secara sembarangan menyalin gen dari satu organisme ke organisme lain kita berisiko mendapat penundaan yang kuat dalam proses penerjemahan. Memang, sementara banyak kodon gen ini jarang ada dalam organisme baru, ribosom akan menunggu lebih lama ketika tRNA yang diinginkan akhirnya tiba.
( ). .Primer yang tumpang tindih disintesis sedemikian rupa sehingga setelah perakitan, gen berukuran penuh terbentuk, pada sisi-sisi di mana terdapat situs pemotongan dengan endonukleas yang sama, lokasi yang terletak di polylinker vektor. Dengan demikian, jika proses endonuklease yang diperlukan (dalam contoh saya adalah enzim-enzim BamH1 dan yang EcoR1 ) vektor dan gen yang dirakit, maka mereka membentuk ujung lengket yang dapat "mengenal satu sama lain" karena interaksi nukleotida komplementer ujung lengket. Tetapi pengenalan saja tidak cukup, karena sejauh ini ujung lengket saling menempel hanya karena ikatan hidrogen yang rapuh. Masalah ini diselesaikan dengan menambahkan campuran reaksi secara bersamaan memotong vektor, memotong gen dan enzim DNA ligase ., yang menghilangkan kerusakan rantai pada molekul DNA untai ganda.Faktor penting lain dalam perakitan gen adalah kenyataan bahwa frekuensi satu atau lain kodon berbeda untuk organisme yang berbeda, sementara di dalam sel biasanya ada lebih banyak tRNA yang sesuai dengan kodon yang lebih "populer". Karena banyak asam amino dikodekan oleh beberapa kodon, sangat mungkin bahwa secara sembarangan menyalin gen dari satu organisme ke organisme lain kita berisiko mendapat penundaan yang kuat dalam proses penerjemahan. Memang, sementara banyak kodon gen ini jarang ada dalam organisme baru, ribosom akan menunggu lebih lama ketika tRNA yang diinginkan akhirnya tiba. , , , , , . E. coli .Sekarang beberapa kata tentang promotor. Pemilihan promotor yang cocok sangat penting, karena proses transkripsi sangat bergantung padanya. Promotor secara kondisional dibagi menjadi kuat, sedang dan lemah. "Kekuatan" promotor ditentukan oleh seberapa aktif gen ditranskripsikan di bawah kendalinya, semua hal lain dianggap sama: semakin transkripsi aktif, semakin kuat promotor. Jelas, ketika kita ingin membuat penghasil protein, kita harus mulai dengan promotor yang kuat. Dalam beberapa kasus, transkripsi yang terlalu cepat (karenanya, terjemahan aktif) membahayakan sel, dalam hal ini Anda dapat mencoba menggunakan promotor yang lebih lemah. Meskipun sebenarnya jauh lebih mudah untuk mempengaruhi aktivitas transkripsi dari produsen yang sudah ada daripada membuat yang baru.Hal lain yang penting. Seringkali, protein yang dikodekan oleh vektor memiliki efek yang sangat negatif pada kelangsungan hidup bakteri. Tidak hanya sintesis protein-protein ini memakan banyak sumber daya (dan jumlah protein target harus baik tidak kurang dari 10% dari total berat kering sel), mereka juga melayang bolak-balik dalam sitoplasma dengan beban mati ! Oleh karena itu, untuk saat ini, lebih baik mematikan ekspresi gen asing ke sel. Untuk tujuan ini, sistem ekspresi terkontrol telah dikembangkan yang memungkinkan Anda untuk "mengaktifkan" ekspresi gen yang kita butuhkan "sesuai perintah." Yang paling umum adalah:
, , , , , . E. coli .Sekarang beberapa kata tentang promotor. Pemilihan promotor yang cocok sangat penting, karena proses transkripsi sangat bergantung padanya. Promotor secara kondisional dibagi menjadi kuat, sedang dan lemah. "Kekuatan" promotor ditentukan oleh seberapa aktif gen ditranskripsikan di bawah kendalinya, semua hal lain dianggap sama: semakin transkripsi aktif, semakin kuat promotor. Jelas, ketika kita ingin membuat penghasil protein, kita harus mulai dengan promotor yang kuat. Dalam beberapa kasus, transkripsi yang terlalu cepat (karenanya, terjemahan aktif) membahayakan sel, dalam hal ini Anda dapat mencoba menggunakan promotor yang lebih lemah. Meskipun sebenarnya jauh lebih mudah untuk mempengaruhi aktivitas transkripsi dari produsen yang sudah ada daripada membuat yang baru.Hal lain yang penting. Seringkali, protein yang dikodekan oleh vektor memiliki efek yang sangat negatif pada kelangsungan hidup bakteri. Tidak hanya sintesis protein-protein ini memakan banyak sumber daya (dan jumlah protein target harus baik tidak kurang dari 10% dari total berat kering sel), mereka juga melayang bolak-balik dalam sitoplasma dengan beban mati ! Oleh karena itu, untuk saat ini, lebih baik mematikan ekspresi gen asing ke sel. Untuk tujuan ini, sistem ekspresi terkontrol telah dikembangkan yang memungkinkan Anda untuk "mengaktifkan" ekspresi gen yang kita butuhkan "sesuai perintah." Yang paling umum adalah:- Sebuah sistem yang didasarkan pada elemen pengatur lactose operon E. coli ( lac -peron) dan promotor yang kuat.
Faktanya adalah E. coli memiliki aturan gizi sendiri. Pertama, ada mekanisme untuk menekan aktivitas lac -peron, yang diaktifkan hanya ketika laktosa tidak memasuki sel. Ini logis: mengapa membuang energi untuk sintesis apa yang tidak berguna? Tetapi begitu laktosa mulai memasuki sel dalam jumlah yang cukup, mekanisme ini dimatikan.
Namun, ada mekanisme kedua untuk menekan aktivitas lac -peron. Jika ada glukosa dalam medium, maka sel makan secara eksklusif pada glukosa, karena mengaktifkan mekanisme kedua penghambatan transkripsi lac- peron. Jadi, lac -peron hanya aktif ketika hanya ada laktosa di ruang sekitar sel. Kekurangan dari operon laktosa adalah promotor yang sangat lemah, oleh karena itu, pada strain produsen digantikan oleh yang kuat. Promotor yang kuat sering berasal dari patogen. Promotor terkuat yang paling banyak digunakan dalam rekayasa genetika prokariota diisolasi dari virus bakteri - bakteriofag . Misalnya, promotor fag T7 banyak digunakan.
Ngomong-ngomong, beberapa promotor kuat untuk rekayasa genetika tanaman juga diisolasi dari virus, misalnya, ini adalah promotor virus mosaik kembang kol.

Seperti disebutkan di atas, E. coli tidak memiliki RNA polimerase yang mengenali promotor bakteriofag, oleh karena itu, gen RNA polimerase dari bakteriofag yang sesuai sebelumnya dimasukkan ke dalam produsen.
Sistem sintesis protein E. coli yang populer membawa gen T7 fag RNA polimerase di bawah kendali promotor RNA polimerase bakteri, diatur oleh mekanisme lakon. Jika strain ini ditransformasikan dengan vektor yang membawa gen target di bawah kendali "regulasi promotor T7 fag + dari kompleks tipe promotor", maka mekanisme dua tingkat menghambat transkripsi gen target akan muncul.
Saat menggunakan desain ini, glukosa dan laktosa ditambahkan ke media nutrisi secara bersamaan. Untuk beberapa waktu, sel-sel akan memakan glukosa dan membelah diri dengan tenang, karena sintesis protein asing ditekan sepenuhnya. Pada saat glukosa berakhir dan sel-sel beralih ke metabolisme laktosa, sudah akan ada cukup biomassa dalam kultur, itu hanya saat untuk memulai sintesis protein yang kita butuhkan. Prosedur ini disebut "induksi otomatis."
Anda dapat melakukannya dengan cara lain: jangan menambahkan glukosa dan laktosa ke media nutrisi, dan kemudian, ketika biakan mencapai kepadatan yang diinginkan, tambahkan apa yang akan diambil sel untuk laktosa, tetapi tidak dapat memetabolisme atau menghancurkannya. Sekarang, IPTG digunakan sebagai induktor.
- Suatu sistem yang didasarkan pada mekanisme pengaturan promotor pL dari bakteriofag λ .
Promotor ini dinonaktifkan oleh protein penekan cI. Dalam hal ini, bentuk termosensitif dari protein yang disebut cI857 ini ditemukan: faktor transkripsi ini mempertahankan fungsi pada suhu sekitar 30 ° C dan kehilangannya pada 42 ° C. Oleh karena itu, ketika menggunakan sistem seperti itu, kultur bakteri pertama-tama ditumbuhkan dengan kepadatan yang diinginkan pada 30 ° C, dan kemudian suhu dinaikkan menjadi 42 ° C, sehingga memulai sintesis protein target.
Nah, vektor dirancang. Maka hal kecil adalah menemukan metode yang cocok untuk dimasukkan ke dalam sel bakteri. Tetapi ini adalah kisah yang sangat berbeda.