Dalam publikasi saya sebelumnya , dua pertanyaan dipertimbangkan: dasar-dasar biologi molekuler dan metode untuk membuat konstruksi yang kita butuhkan untuk gen berdasarkan vektor plasmid. Sekarang Anda perlu mencari cara bagaimana memperkenalkan vektor plasmid ke dalam sel bakteri, yaitu, untuk membuat "transformasi". Pada saat yang sama, kita akan belajar sesuatu tentang struktur membran bakteri dan bagaimana cara mengatasinya, serta mengapa beberapa bakteri disebut kompeten, bagaimana semua ini terkait dengan racun bakteri dan ketahanan bakteri terhadap antibiotik, dan banyak fakta menarik lainnya.
Jadi, kita memiliki vektor plasmid di tangan kita dan ada strain bakteri yang cocok untuk vektor ini, tetap memperkenalkan vektor ke dalam bakteri.
Dalam
artikel sebelumnya ,
karya Frederick Griffith disebutkan, yang dengan jelas menunjukkan fakta bahwa DNA dapat secara spontan menembus sel bakteri. Kemampuan sel bakteri untuk menyerap molekul DNA lingkungan disebut "kompetensi" (yang menimbulkan banyak lelucon tentang fakta bahwa bahkan uniseluler in vitro yang kompeten, dan rekan laboratorium Anda tidak). Jika penyerapan tidak terjadi di bawah pengaruh beberapa kondisi yang diciptakan secara artifisial, maka kita berbicara tentang "kompetensi alami". Bagaimana transportasi DNA serupa di dalam bakteri terjadi?
1) Struktur membran sel bakteri
Untuk mulai dengan, kami secara singkat mempertimbangkan struktur membran bakteri gram positif dan gram negatif.
Cangkang bakteri gram negatif terdiri dari dua lapisan lipid: membran luar dan membran dalam (bagian dalam juga disebut plasma). Di antara mereka adalah ruang periplasmik yang diisi dengan protein dan peptidoglikan melakukan fungsi-fungsi penting.
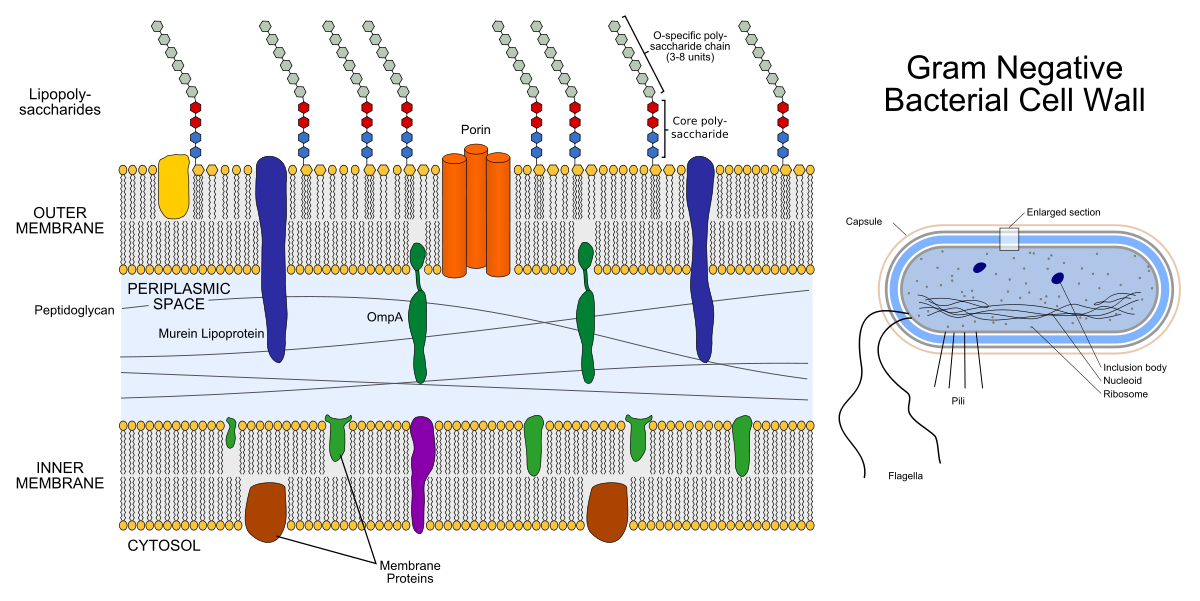 Di sebelah kiri adalah struktur membran bakteri gram negatif. Gizmos berwarna di dalam membran adalah protein membran, dan rantai yang mencuat adalah lipopolisakarida. Di sebelah kanan adalah struktur bakteri gram negatif itu sendiri.
Di sebelah kiri adalah struktur membran bakteri gram negatif. Gizmos berwarna di dalam membran adalah protein membran, dan rantai yang mencuat adalah lipopolisakarida. Di sebelah kanan adalah struktur bakteri gram negatif itu sendiri.Peptidoglikan adalah heteropolimer yang terdiri dari N-acetylglucosamine (NAG) dan N-acetylmuramic acid (NAM) yang berikatan silang secara kovalen. Ternyata rantai ....- NAG-NAM-NAG-NAM-NAG- .... "Hetero" dalam kata "heteropolymer" berarti bahwa polimer tidak terdiri dari elemen yang sama (seperti polietilen), tetapi dari dua yang berbeda. Enzim sintesis peptidoglikan, transpeptidase, adalah target untuk antibiotik β-laktam (antibiotik penisilin dan sefalosporin): mereka menghambat aktivitasnya, sebagai akibat dari proses pembelahan bakteri, tidak ada cukup peptidoglikan untuk kedua sel anak. Lapisan peptidoglikan juga merupakan target untuk enzim lisozim: ia menghancurkan ikatan kovalen antara NAG dan NAM. Dalam kedokteran, lisozim digunakan sebagai antiseptik, misalnya, ia merupakan komponen aktif dari obat-obatan tertentu untuk pengobatan sakit tenggorokan, dan dalam industri makanan dapat digunakan sebagai pengawet (suplemen makanan E1105 - ini adalah lisozim).
Peptidoglikan. Gambar tersebut menunjukkan bagian yang sesuai dengan satu NAG (itu di sebelah kiri) dan satu NAM (di sebelah kanan), yang saling berhubungan melalui atom oksigen. (Bagi mereka yang tidak menyukai kebebasan kimia seperti ini: "ikatan melalui atom oksigen" ini disebut β- (1,4) -glycoside).Amplop bakteri gram positif hanya terdiri dari membran internal (plasma), ruang periplasmik yang sangat tipis dan lapisan peptidoglikan eksternal yang lebih tebal dari pada bakteri gram negatif.
Rintangan yang menghalangi setiap senyawa dalam sitoplasma bakteri tidak dapat diatasi untuk semua hal kecuali air, serta molekul-molekul kecil non-polar dan hidrofob (molekul oksigen dan nitrogen, karbon dioksida dan lain-lain). Penetrasi senyawa-senyawa ini melalui kulit disebut "transpor pasif" ("difusi sederhana"), selalu terjadi di sepanjang gradien konsentrasi. Molekul non-polar dapat dengan bebas melewati membran karena volume internal membran juga non-polar (di dalam membran mereka terdiri dari residu asam lemak hidrofobik), dan senyawa non-polar sangat larut dalam pelarut non-polar (alkohol, aseton, asam lemak).
Sebuah gambar komik di mana hidrofobisitas (hidrofobia) residu asam lemak di dalam membran dua lapis dimainkan.Pada gilirannya, molekul polar besar (asam amino, monosakarida, nukleotida), partikel bermuatan (ion) dan makromolekul (DNA, protein) tidak dapat menembus melalui membran sel begitu saja, karena mereka larut hanya dalam pelarut polar (misalnya, dalam air) . Lalu bagaimana molekul DNA masuk ke dalam sel bakteri?
2) Mekanisme pengangkutan senyawa yang diinduksi ke dalam sel dari lingkungan
Pertama-tama, kita memiliki fakta nyata bahwa bakteri tidak dapat hidup hanya dengan air, oksigen, dan karbon dioksida, karena untuk kehidupan diperlukan banyak senyawa lain. Jadi ada cara untuk mengirimkan senyawa ini ke dalam sel. Metode tersebut adalah "difusi difasilitasi" dan "transportasi aktif".
2.1) Difusi cahaya
Contoh difusi yang difasilitasi (meskipun kasus khusus ini, sebaliknya, berbahaya bagi bakteri) adalah pengangkutan ion kalium melalui membran sel bakteri dalam kombinasi dengan valinomycin, antibiotik yang disintesis oleh beberapa jamur. Valinomisin bebas tidak dapat menembus ke dalam sel, karena kelompok kutub terpapar secara eksternal. Ion kalium juga merupakan "jalur tertutup".
 Patung yang menggambarkan struktur tiga dimensi kompleks valinomisin dan ion kalium (ion digambarkan sebagai bola di tengah). Itu dipasang di depan pintu masuk ke M. M. Shemyakin dan Yu A. A. Ovchinnikov Institut Kimia Bioorganik, Akademi Ilmu Pengetahuan Rusia (Moskow), di mana mekanisme aktivitas antibakteri valinomycin dan struktur tiga dimensi kompleksnya dengan ion kalium didirikan.
Patung yang menggambarkan struktur tiga dimensi kompleks valinomisin dan ion kalium (ion digambarkan sebagai bola di tengah). Itu dipasang di depan pintu masuk ke M. M. Shemyakin dan Yu A. A. Ovchinnikov Institut Kimia Bioorganik, Akademi Ilmu Pengetahuan Rusia (Moskow), di mana mekanisme aktivitas antibakteri valinomycin dan struktur tiga dimensi kompleksnya dengan ion kalium didirikan.Tetapi setelah pembentukan kompleks valinomisin dengan ion kalium, ia mengubah struktur tiga dimensi sehingga seluruh permukaan luarnya menjadi hidrofob (non-polar), dan semua gugus hidrofilik tersembunyi di dalamnya. Dalam hal ini, ion kalium disembunyikan di dalam molekul valinomisin, oleh karena itu, kompleks valinomisin + ion kalium dapat menembus melalui membran. Kompleks ini dapat terurai menjadi valinomisin dan ion bebas kapan saja baik di dalam sel maupun di luar, tetapi difusi ion secara umum memfasilitasi penyamaan konsentrasi mereka baik dalam ruang intraseluler dan ekstraseluler, dan sel mengontrol pengangkutan ion karena suatu alasan: konsentrasi tinggi mereka fatal. Di sinilah efek antibakteri valinomisin didasarkan.
Representasi skematik antibiotik valinomisin jamur yang dikomplekskan dengan ion kalium.Contoh lain adalah pori-pori yang tidak terkontrol, yang merupakan protein transmembran besar yang memungkinkan zat-zat tertentu untuk secara bebas melewati membran sesuai dengan gradien. Contoh dari pori transmembran seperti itu adalah
Staphylococcus aureus Staphylococcus aureus α-toksin (nama lain untuk toksin tersebut adalah α-hemolysin. Ini terdiri dari bagian "hemo" dan "lisin". "Hemo" berarti sel darah merah; "lisin" berarti sel darah merah; dari kata "lisis", yaitu, "kematian sel")
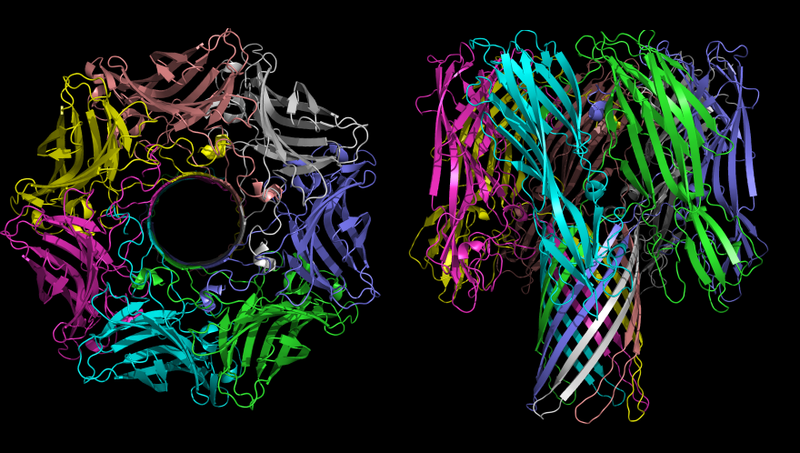 Struktur tiga dimensi oligomer α-hemolisin - toksin dari bakteri Staphylococcus aureus. Pada gambar di sebelah kiri, waktu terlihat jelas.
Struktur tiga dimensi oligomer α-hemolisin - toksin dari bakteri Staphylococcus aureus. Pada gambar di sebelah kiri, waktu terlihat jelas.Fungsi utama α-hemolisin dalam proses patogen adalah pembuatan saluran yang tidak terkontrol dalam membran sel, yang, tergantung pada kondisi lingkungan, dapat melewati ion monovalen, ion kalsium, atau ATP. Seperti disebutkan di atas, ketidakseimbangan ion sangat beracun, dan pengangkutan ATP
dari sel berbahaya, karena ATP merupakan mata rantai yang sangat diperlukan dalam hampir semua proses kimiawi sel apa pun.
2.2) Transportasi aktif
Seperti dapat dilihat dari contoh-contoh di atas, difusi yang difasilitasi tidak memerlukan energi. Sebaliknya, transpor aktif adalah penyerapan atau pelepasan sesuatu oleh suatu sel dengan pengeluaran energi untuk setiap aksi transfer. Ini dapat berupa pengangkutan molekul atau ion polar besar, dan pengangkutan ion dapat dilakukan di sepanjang gradien konsentrasi dan melawannya. Tentu saja, transpor aktif adalah proses terkontrol di mana seluruh rangkaian protein intraseluler dan transmembran terlibat. Jadi, pertama sel perlu menghabiskan sumber daya untuk mensintesis semua protein ini, dan kemudian lagi menghabiskan energi untuk penggunaan sistem transportasi ini.
Contoh transport aktif adalah sistem transportasi yang bergantung pada energi dari senyawa hidrofobik dan amfifilik dari sel kembali ke lingkungan (efflux).
Sistem transpor aktif antibiotik dari sel bakteri ke luar, terdiri dari tiga protein yang membentuk kompleks transmembran tunggal. Gambar tersebut mencerminkan fakta bahwa protein pengangkut eflux (violet) dapat "menangkap" molekul obat baik secara langsung di sitoplasma dan di membran.Sistem ini sangat menarik karena menyebabkan multi-resistensi beberapa bakteri terhadap antibiotik: sel menghilangkannya sebelum mereka benar-benar dapat merusaknya. Selain itu, lebih dari setengah kasus resistensi
multi- obat
Pseudomonas aeruginosa Pseduomonas aeruginosa dikaitkan dengan transportasi aktif antibiotik. Lebih penting lagi, sistem eflux aktif dari antibiotik "baru" telah ditemukan yang digunakan terhadap bakteri yang resisten terhadap obat antibakteri yang biasa digunakan. Misalnya, obat
Linezolid yang digunakan dalam kasus ketidakefektifan antibiotik klasik dilemparkan kembali oleh sistem penghabisan berdasarkan protein AcrAB dan TolC. Fakta lain yang menarik adalah bahwa bakteri pintar tidak membuang energi pada sintesis protein berskala besar dari sistem penghabisan dalam situasi-situasi ketika mereka tidak diperlukan. Tetapi begitu zat-zat berbahaya bagi sel muncul, sintesisnya diaktifkan dengan cepat dan bakteri mulai aktif melawan. Omong-omong, penghabisan bukan satu-satunya mekanisme untuk melindungi bakteri dari antibiotik, saya akan menulis artikel tentang metode lain juga.
Struktur linezolid adalah antibiotik yang digunakan melawan strain resisten bakteri gram positif.3) Kompetensi sel bakteri
Jadi, kami memeriksa mekanisme transportasi transmembran senyawa yang tidak dapat menembus penghalang ini tanpa bantuan dari luar. Dalam kondisi alamiah, pengangkutan DNA ke dalam bakteri dilakukan hanya melalui pengangkutan aktif karena kerja sama seluruh spektrum baik protein intraseluler maupun transmembran. Kemampuan sel yang tak terinduksi untuk menyerap DNA dari luar disebut "kompetensi alami."
3.1) Kompetensi alami sel bakteri
Hipotesis yang paling kuat tentang kemunculan kompetensi alami bakteri adalah asumsi bahwa mekanisme ini memungkinkan bakteri untuk bertahan hidup dalam kondisi ekstrem bagi mereka (kondisi ekstrem untuk sel disebut "stres"). Faktanya adalah bahwa stres sering menyebabkan kerusakan pada "kromosom" DNA bakteri, dan jika sebuah sel ingin bertahan hidup, maka sel itu perlu "memperbaiki" "kromosom" -nya ("perbaikan" seperti itu disebut "perbaikan"). Selain itu, diketahui bahwa jika sekuens DNA dimasukkan ke dalam sel yang mirip dengan beberapa bagian dari DNA sel itu sendiri, maka sangat mungkin untuk mengintegrasikan bagian "baru" ini ke dalam genomnya. Apa yang terjadi dalam kondisi stres? Beberapa sel mati, menjadi donor DNA bagi saudara-saudara mereka yang masih berjuang untuk hidup. Memang, dalam kultur bakteri yang sama, sangat mungkin bahwa genom dari perwakilan sangat mirip, sehingga bakteri yang masih hidup dapat menggunakan DNA kerabat yang mati untuk memperbaiki genom mereka.
Selain itu, di bawah kondisi yang nyaman dalam kultur sel, hanya sebagian kecil dari mereka yang memiliki kompetensi alami, dan proporsi sel yang kompeten dari jumlah totalnya adalah yang terbesar, kemudian kultur tersebut berada dalam fase pertumbuhan tertentu - fase
log .
 Sebidang tipikal bakteri dalam biakan seiring waktu. Fase log adalah fase pertumbuhan, diikuti oleh dataran tinggi dan tahap kepunahan, di mana sel-sel sudah kekurangan nutrisi dan racun menumpuk.
Sebidang tipikal bakteri dalam biakan seiring waktu. Fase log adalah fase pertumbuhan, diikuti oleh dataran tinggi dan tahap kepunahan, di mana sel-sel sudah kekurangan nutrisi dan racun menumpuk.Tentu saja, peneliti ingin membuat proses transfer vektor plasmid ke dalam sel seefisien mungkin: semakin efisien proses transformasi, semakin sedikit vektor plasmid yang dibutuhkan untuk menjamin hasil yang baik. Oleh karena itu, dalam kondisi laboratorium, mereka biasanya tidak bergantung pada sel itu sendiri, melainkan membuat sel dengan kompetensi buatan.
3.2) Kompetensi buatan sel bakteri
Ada dua metode utama untuk menciptakan kompetensi buatan bakteri: pengobatan kultur sel dengan ion divalen diikuti oleh pemanasan jangka pendek dan yang disebut elektroporasi.
3.2.1) Perawatan kultur sel dengan ion divalen, diikuti dengan pemanasan jangka pendek.
Biasanya, di laboratorium ada lemari es dengan suhu sekitar -80 derajat Celcius (kadang-kadang disebut "kelvinators" untuk menghormati salah satu perusahaan manufaktur di negara kita), yang tersumbat dengan tabung reaksi dengan sel-sel dalam fase
log . Hal ini dilakukan agar tidak membuang waktu membuat mereka setiap kali dibutuhkan (dan itu membutuhkan banyak waktu, itu tidak akan bekerja lebih cepat daripada dalam sehari). Dan agar selama penyimpanan sel tidak mati karena pembekuan, cryoprotectants ditambahkan ke medium: gliserol, propilen glikol atau dimetil sulfoksida.
Lemari es laboratorium dalam pandangan penulis gambar.Ketika seorang peneliti perlu mengubah strain tertentu, ia mengangkat sel-sel dari kelvinator dan menempatkan tabung di dalam es. Kemudian, kation divalen dan vektor plasmid yang kita butuhkan ditambahkan ke sel-sel dalam fase pertumbuhan
log . Tabung reaksi tidak boleh meninggalkan es. Donor tipikal dari kation semacam itu adalah kalsium klorida (CaCl2), dalam hal ini, kation divalen kami adalah Ca2 +. Tidak ada penjelasan tegas tentang bagaimana Ca2 + membantu DNA untuk menembus bakteri:
- Beberapa sumber mengklaim bahwa konsentrasi Ca2 + yang tinggi menyebabkan celah pada membran sel dan kerusakan DNA, dan sebelumnya kami menemukan bahwa kerusakan DNA adalah sinyal bagi sel untuk menjadi kompeten;
- Menurut sumber lain, ion ini berfungsi sebagai "jembatan" yang menempelkan vektor plasmid ke sel. Faktanya adalah ion bermuatan positif menempel secara simultan pada kelompok polisakarida bermuatan negatif pada membran luar bakteri dan pada kelompok DNA plasmid bermuatan negatif. Jika tidak ada ion dalam medium, maka DNA akan diusir dari sel, dan ini sama sekali tidak berguna bagi peneliti.
Ada kemungkinan bahwa kedua hipotesis itu benar.
Seharusnya mekanisme partisipasi ion Ca2 + dalam transformasi. Pertama, ion menempel pada kelompok DNA bermuatan negatif (lingkaran kuning) dan polisakarida yang berlabuh di membran (lingkaran kuning). Kemudian, karena ion, semacam jembatan terbentuk: "polisakarida (-)" - "ion Ca (2+)" - "DNA (-)", yang memperbaiki DNA pada permukaan sel.Setelah inkubasi bakteri pada suhu mendekati nol, yang disebut sengatan panas dihasilkan - pemanasan jangka pendek kultur hingga 42 derajat. Biasanya, durasi perlakuan panas adalah 30-45 detik, setelah itu tabung reaksi dengan bakteri kembali ke penangas es (dengan kejutan panas yang lebih lama sel-sel dapat mati). Alasan yang mungkin untuk keefektifan syok panas adalah mekanisme yang sama: suhu tinggi dapat merusak DNA dan protein (ini menginduksi kompetensi), dan juga dapat menciptakan "lubang" di membran sel yang dilalui oleh vektor plasmid yang menempel pada membran melalui ion kalsium yang menembus.
Kemudian sel-sel dilapisi pada cawan Petri dengan media selektif dan koloni tumbuh di atasnya.
3.2.2) Elektroporasi
Dasar dari metode
elektroporasi adalah kenyataan bahwa medan listrik dapat merusak membran sel. Artinya, secara harfiah, elektroporasi adalah penciptaan pori-pori oleh medan listrik.
Pertama, bakteri dan vektor plasmid ditempatkan dalam tabung khusus (kuvet). Kemudian cuvette ditempatkan dalam perangkat khusus, yang untuk sepersekian detik menciptakan perbedaan potensial 0,1-10 kV, yang pada gilirannya menyebabkan kerusakan reversibel pada membran dan vektor dapat dengan bebas menembus ke dalam sel. Setelah beberapa waktu, kultur bakteri diunggulkan di cawan Petri dengan media selektif.
Persyaratan penting untuk media cair di mana sel dan vektor terletak pada saat elektroporasi adalah konsentrasi ion yang rendah: jika tidak, korsleting dapat terjadi.
Kata penutup
Setiap koloni di piring adalah keturunan pengembangbiakan
satu bakteri tunggal (yaitu, jika kita melihat 100 koloni di piring, itu berarti bahwa pada awalnya ada 100 bakteri yang hidup, yang kemudian mulai membelah).
Setelah koloni muncul di piring, peneliti memilih salah satu koloni dan mentransfer sel dari koloni ke media nutrisi cair selektif baru. Setelah kultur sel baru "tumbuh" sekali lagi ditanam di cangkir baru dengan media nutrisi selektif yang sama. Sebagai hasilnya, kita mendapatkan cangkir di mana hanya keturunan bakteri dari satu koloni yang tumbuh (yaitu, pada kenyataannya mereka semua adalah klon dari satu bakteri tunggal). Dan hanya sel-sel ini yang akan digunakan di masa depan untuk mendapatkan protein yang kita butuhkan dalam volume besar medium nutrisi selektif cair.Baiklah, mari rangkum hasil antara:- Kami mensintesis gen yang kami butuhkan;
- Masukkan ke dalam vektor plasmid yang cocok;
- Strain produsen yang cocok dipilih dan ditransformasikan;
- Punya budaya klon;
- Akhirnya, kami memindahkan sebagian dari klon-klon ini ke volume besar medium nutrisi untuk mendapatkan protein yang kami butuhkan dalam volume besar.
Langkah selanjutnya: untuk mengisolasi protein dari biomassa yang dihasilkan dalam bentuk semurni mungkin. Tetapi ini adalah kisah yang sangat berbeda.