Pertanyaan tentang alat penglihatan adalah bagian nyata dari neurobiologi. Volume besar literatur dan empat hadiah Nobel dikhususkan untuk masalah ini, tetapi dalam situasi ini orang tidak bisa tidak memperhatikan bahwa perangkat penglihatan mamalia yang dijelaskan dalam buku-buku teks tidak mengatasi tugas tersebut. Tujuan dari esai ini adalah untuk menunjukkan serangkaian alasan mengapa Anda tidak menutup mata terhadap hal ini. Bahkan, potret rahasia penglihatan akan disajikan, mulai dari berbagai detail kecil di awal aliran informasi visual pada mamalia, ancaman mengabaikan mereka, dan diakhiri dengan tumpukan masalah dalam memahami pemrosesan otak di ujung jalan.
Perangkat sistem visi
Di mata buku teks tentang penglihatan, kita melihat dalam tiga tahap. Tahap pertama: cahaya memasuki retina dan diubah menjadi eksitasi saraf fotoreseptor - neuron sensorik retina. Selain itu, mata menormalkan kontras dan kecerahan, memfokuskan gambar.
Tahap kedua: proses di retina, ketika peta rangsangan saraf fotoreseptor ditransformasikan menjadi aliran informasi paralel, masing-masing elemen yang mengkodekan aspek informasi visualnya sendiri. Sel-sel ganglion, yang aksonnya membentuk saraf optik, di retina jauh lebih kecil daripada fotoreseptor, kompresi informasi pertama kali terjadi. Satu sel ganglion mungkin bertanggung jawab untuk mengkodekan sinyal dari beberapa di pusat menjadi ratusan dan ribuan fotoreseptor pada pinggiran retina. (Gambar 1) Sel-sel ganglion dibagi menjadi dua jenis utama, tergantung pada rute mana dari saraf optik, sinyal memasuki korteks dari belahan otak. Satu jenis yang berlaku secara kuantitatif (80% pada orang) memberikan ketajaman visual yang tinggi dan persepsi warna, kontras gambar yang stabil penting bagi mereka. Untuk alasan historis, mereka disebut sel PC atau sel kerdil, karena sinyal dari mereka melewati lapisan parvocellular (sel kecil) dari badan engkol lateral. Tipe kedua dari sel MC, sinyalnya melewati lapisan magnocellular (sel besar) dari badan engkol lateral. Mereka bertanggung jawab atas persepsi gerakan, karena mereka memiliki sensitivitas yang meningkat terhadap gambar yang berkedip atau bergerak.


 Gambar 1. Mengukur sifat-sifat bidang reseptif. (A) Respons neuron (urutan potensial aksi atau "adhesi") dikendalikan oleh rekaman ekstraseluler hewan yang dianestesi oleh elektroda di badan perantara crankshaft intermediate (LGN). Stimuli dikirim ke bagian bidang visual yang sesuai dengan neuron. (B) Tabel respons bersyarat dari berbagai jenis neuron. Setiap kolom menunjukkan respons dari satu jenis neuron. Setiap baris menunjukkan respons terhadap satu jenis stimulus. Misalnya, neuron yang selektif dalam arah gerakan (kolom kedua) hanya bereaksi ketika stimulus bergerak melalui bidang reseptif dari kiri ke kanan. Neuron yang memiliki bidang reseptor standar "pusat / lingkungan" toleran terhadap ukuran dan motilitas stimulus. Jenis neuron yang tidak standar menunjukkan persyaratan stimulus yang jauh lebih tanpa kompromi.
Gambar 1. Mengukur sifat-sifat bidang reseptif. (A) Respons neuron (urutan potensial aksi atau "adhesi") dikendalikan oleh rekaman ekstraseluler hewan yang dianestesi oleh elektroda di badan perantara crankshaft intermediate (LGN). Stimuli dikirim ke bagian bidang visual yang sesuai dengan neuron. (B) Tabel respons bersyarat dari berbagai jenis neuron. Setiap kolom menunjukkan respons dari satu jenis neuron. Setiap baris menunjukkan respons terhadap satu jenis stimulus. Misalnya, neuron yang selektif dalam arah gerakan (kolom kedua) hanya bereaksi ketika stimulus bergerak melalui bidang reseptif dari kiri ke kanan. Neuron yang memiliki bidang reseptor standar "pusat / lingkungan" toleran terhadap ukuran dan motilitas stimulus. Jenis neuron yang tidak standar menunjukkan persyaratan stimulus yang jauh lebih tanpa kompromi.Kedua jenis sel ini memiliki jenis organisasi pusat / lingkungan: strategi pengkodean yang konon universal untuk informasi visual. (Gambar 2) Stimulus terbaik untuk mencapai respons paling energik dari sel-sel ganglion adalah lebih atau kurang bulat dan kecil pada retina, dan semakin intensif menonjol terhadap latar belakang sekitarnya (titik terang atau gelap), semakin energik adalah respons sel.
Tahap ketiga: sinyal diproses di otak. Dari sudut pandang yang diterima secara umum, proses utama pemrosesan otak dimulai pada korteks visual primer. Pengodean ulang lain yang signifikan terjadi, neuron tidak hanya merespons bintik-bintik gelap dan terang pada permukaan retina, tetapi juga menjadi selektif sensitif terhadap batas-batas kontras yang berorientasi, bagian garis yang berorientasi dan ujungnya. Ada konfigurasi rangsangan yang lebih kompleks, ketika neuron merespons dengan tepat pada batas kontras tanpa mempedulikan posisi, jika saja ia berada dalam zona tanggung jawab sel. Pemrosesan "kompleks" dalam korteks visual ini digunakan sebagai tingkat kedua abstraksi dalam persepsi gambar statis, di mana pengakuan telah mencapai tingkat persepsi batas yang lebih umum tanpa memandang posisi mereka. Akibatnya: pertama, retina mentransmisikan sinyal sederhana ke korteks serebral; kedua, neuron dari korteks visual primer menggunakan sinyal sederhana ini untuk mendeteksi batas dan garis yang kontras; dan ketiga, blok bangunan fundamental ini digunakan untuk menentukan batas-batas seluruh objek dan menciptakan persepsi visual.
Mungkin tidak ada yang berasumsi bahwa sudut pandang ini akan berubah menjadi dogma fundamental granit. Deteksi tepi telah dilihat sebagai satu-satunya cara bagi mamalia untuk melihat bagaimana evolusi menemukan solusi sempurna untuk masalah persepsi gambar yang efektif. Dalam esai ini, sudut pandang ini akan dipertanyakan dan pertanyaan tidak nyaman akan ditanyakan tentang sifat visi. Mereka telah dibesarkan lebih dari sekali dalam sejarah studi tentang penglihatan dan tidak menyarankan jawaban sederhana. Jadi sekarang tugas utama adalah berkonsentrasi pada masalah-masalah ini, menyatukan alasan-alasan keraguan dalam pemahaman modern tentang alat penglihatan.
Retina semua mamalia mengirimkan sinyal yang tidak standar ke otak
Sebuah studi menyeluruh tentang jalur aliran visual dari mata ke korteks serebral pada primata dengan tegas menegaskan bahwa, seperti pada spesies mamalia lainnya, banyak saluran paralel memberikan kontribusi fungsional terhadap aliran visual primata [1-4]. (Gambar 2) Beberapa saluran ini bahkan mengabaikan korteks visual primer dan segera pergi ke departemen hierarki yang lebih tinggi [5]. Banyak dari ini disebutkan dalam studi klasik tentang anatomi kucing dan monyet [6-10], tetapi diabaikan oleh para perintis neurobiologi dalam studi korteks visual. Penggunaan agresif dari prinsip Occam razor memungkinkan untuk memberi makna pada sifat-sifat neuron kortikal, karena mereka membangun fungsinya mengenali garis atau batas semata-mata berdasarkan input dari sel ganglion dengan organisasi "pusat / lingkungan" [11]. Sel-sel ganglion dengan organisasi non-standar dibuang, memiliki jumlah primata yang relatif rendah di retina. Apa peran mereka dalam persepsi visual akan dijelaskan di bawah ini, dan dari sudut pandang para praktisi neurobiologi perlu dicatat bahwa dengan pengenalan elektroda ke dalam saraf optik secara sewenang-wenang, kemungkinan menemukan akson sel ganglion non-standar cukup kecil. Karena sel non-standar jarang terjadi, sebagian besar peneliti memilih untuk mengikuti jalur yang paling tidak resistan, menciptakan metode untuk memproyeksikan rangsangan visual ke retina yang paling cocok untuk mempelajari sel pusat / lingkungan, yang mengubah sel non-standar menjadi "ledakan" statistik dalam data eksperimen.
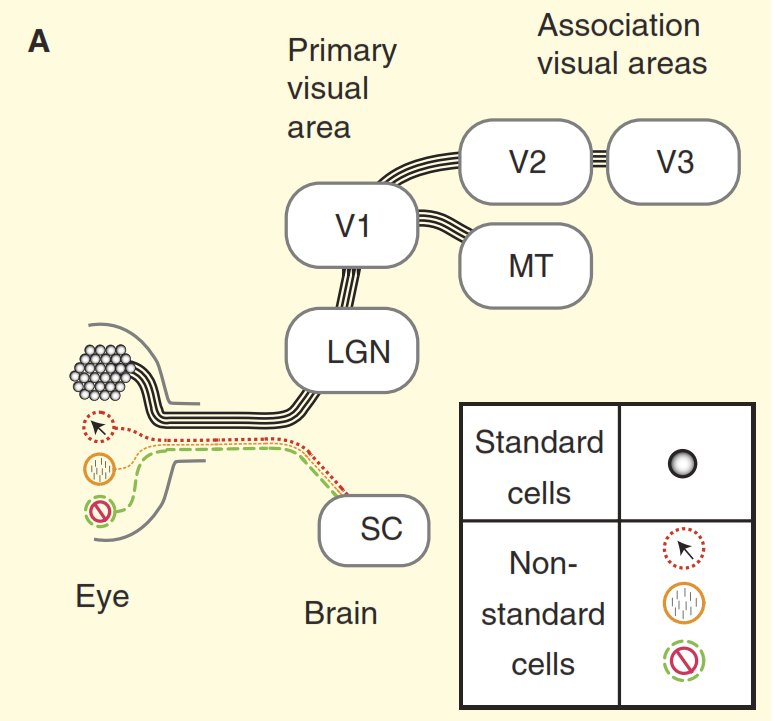
 Gambar 2. Persepsi sistem visual.
Gambar 2. Persepsi sistem visual.
(A) Sudut pandang dari buku pelajaran tentang desain sistem visual primata dengan contoh kera. Retina diisi dengan sejumlah besar sel ganglion standar, yang aksonnya membentuk saraf optik. Retina juga mengandung sejumlah kecil sel ganglion non-standar yang merespons cahaya dengan cara lain, tetapi secara tradisional diasumsikan bahwa mereka diproyeksikan hanya ke pusat otonom otak tengah, yaitu superior dicollis (SC). Sel-sel konsentris menonjol ke inti relay di thalamus, badan engkol perantara (LGN), yang pada gilirannya mentransmisikan sinyal ke korteks visual primer (V1). Dan sel standar melalui intermediate cranked body (LGN) mentransmisikan sinyal ke korteks visual primer (V1) dan kemudian ke bagian yang lebih tinggi dari korteks visual (V2, V3, MT).
(B) Pandangan yang lebih realistis tentang sistem visual primata. Beberapa sel ganglion non-standar meracuni sinyal langsung ke badan engkol perantara (LGN), dan kemudian ke korteks visual primer. Selain itu, dioda atas bukan arah buntu, dari sana sinyal menuju ke badan engkol perantara (LGN) dan seterusnya. Tapi itu tidak semua, dimulai dengan intermediate cranked body (LGN), aliran visual berhenti menjadi searah, dan komponen kebalikannya lebih kuat daripada yang langsung.Masalah kedua diilustrasikan pada Gambar 3. Grafik atas dari gambar menunjukkan respon dari sel "pusat / lingkungan". Skema reaksi dasar sangat familier bagi penglihatan neurobiologis: sel ganglion pusat / lingkungan standar menunjukkan kurva garis respons terhadap frekuensi spasial - respons maksimum terjadi ketika pita kisi tepat bertepatan lebar dengan bagian tengah bidang reseptif pusat / sel lingkungan. " Tetapi sekarang perhatikan respons salah satu sel non-standar dalam saraf optik yang diperoleh dalam percobaan yang sama (Gambar 3b). Kali ini, sel tersebut termasuk jenis sel ganglion konioseluler, satu set komposit tipe sel non-standar, yang biasanya digeneralisasi, jika sama sekali, disebut sebagai "jalur biru". Sel ini memiliki organisasi tipe "penindasan dengan kontras" [6,7,12,13], dalam percobaan ini responsnya hampir sepenuhnya berlawanan dengan respons sel "pusat / lingkungan" standar. Ketika menyesuaikan perbedaan ketajaman visual monyet dan kucing dipertimbangkan, sel-sel ganglion dari “penindasan dengan kontras” berperilaku identik [12,13].
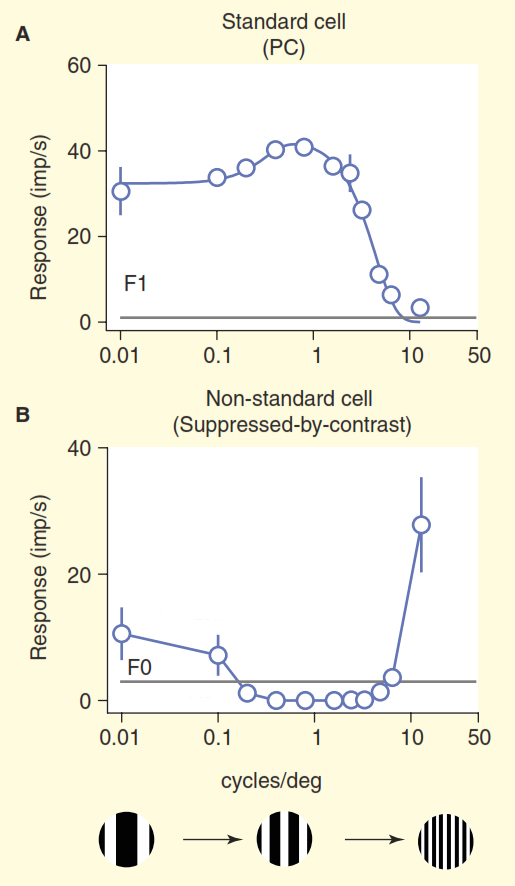 Gambar 3. Perbandingan bidang reseptif standar dan non-standar. Sel dirangsang oleh kisi yang hanyut dengan frekuensi spasial variabel (frekuensi di sini dinyatakan sebagai total lebar dua pita hitam dan putih dalam kisi per derajat sudut pandang). Sel ganglion "pusat / lingkungan" standar (A) berperilaku seperti yang diharapkan: ketika lebar pita kisi dekat dengan lebar tengah bidang reseptif, sel memberikan respons yang kuat. Sel non-standar dengan "penindasan kontras" (B) diam dan bukannya terangsang, selama ia dapat membedakan antara garis-garis hitam dan putih.
Gambar 3. Perbandingan bidang reseptif standar dan non-standar. Sel dirangsang oleh kisi yang hanyut dengan frekuensi spasial variabel (frekuensi di sini dinyatakan sebagai total lebar dua pita hitam dan putih dalam kisi per derajat sudut pandang). Sel ganglion "pusat / lingkungan" standar (A) berperilaku seperti yang diharapkan: ketika lebar pita kisi dekat dengan lebar tengah bidang reseptif, sel memberikan respons yang kuat. Sel non-standar dengan "penindasan kontras" (B) diam dan bukannya terangsang, selama ia dapat membedakan antara garis-garis hitam dan putih.Kita hanya dapat mengasumsikan bahwa sel-sel ini mengenali isian yang monoton dalam bidang tanggung jawabnya, misalnya, langit biru yang jernih, karena ketika lebar batang putih dan hitam kisi berkurang di bawah resolusi mata manusia, mereka bergabung menjadi abu-abu solid. Pemahaman yang benar tentang pengkodean informasi visual untuk sel-sel ini tidak disajikan dalam model visi standar.
Semua non-primata melihat dengan baik menggunakan sel-sel non-standar
Pendukung model penglihatan yang diterima ketika mempertimbangkan primata dapat merujuk pada sejumlah kecil sel non-standar, tetapi ini tidak berlaku untuk retina kelinci, kucing, tikus, dan tikus [7,8,14-16]. Selain itu, dalam jumlah absolut (sekitar 100.000), jumlah sel yang tidak standar dalam retina primata dekat dengan jumlah total sel dalam retina tikus atau kucing [17]. Ketajaman visual pada kucing, kelinci, tikus dan tikus lebih rendah dari pada primata, tetapi ini tidak semuanya. Visi semua makhluk ini memungkinkan Anda untuk berhasil bertahan hidup di alam liar. Siapa pun yang meragukan hal ini diundang untuk mencoba menyelinap pada kelinci di lapangan atau menangkap tikus tanpa menggunakan perangkap tikus (tugas sederhana untuk kucing biasa).
 Gambar 4. Persepsi visual dengan detail rendah.
Gambar 4. Persepsi visual dengan detail rendah.
Pengamat yang akrab dengan orang-orang dalam gambar mengenalinya meskipun detailnya rendah. Ketika mengaburkan gambar, informasi visual yang berguna tetap cukup untuk seseorang. Dari kiri ke kanan: Pangeran Charles, Woody Allen, Bill Clinton, Saddam Hussein, Richard Nixon, Putri Diana.Jelas, jika hanya karena alasan medis, mengapa ahli saraf telah berkonsentrasi pada retina primata. Tetapi konsentrasi selektif seperti itu ketika mencoba untuk memahami visi mengarah pada fakta bahwa para peneliti membawa dan mentransfer hasil persepsi diri ke interpretasi hasil penelitian. Kemasan padat sel-sel ganglion primata sangat cocok untuk mendeteksi buah yang dapat dimakan di pohon, atau untuk membaca edisi terbaru The New York Times, tetapi tidak ada alasan khusus untuk mempertimbangkan tugas-tugas seperti sumum bonum (lat:: kebaikan tertinggi) dari penglihatan. Dalam gambar evolusi umum, ketajaman visual yang sangat tinggi ini dapat dianggap sebagai adaptasi niche khusus untuk mendeteksi objek kontras tinggi pada jarak yang sangat jauh. Kebanyakan mamalia di retina tidak memiliki fovea (fossa pusat di retina) - area khusus dengan ketajaman visual tinggi yang ditemukan pada primata - tetapi penglihatan tanpa sejuta sel ganglion yang padat tetap berfungsi penuh. Buktinya diilustrasikan pada Gambar 4, orang dengan mudah mampu mengenali tanpa ketajaman visual yang biasa [18]. Pada mamalia non-primata, pengkodean rangsangan visual telah dipelajari dengan baik pada kelinci [19-23]. Kelinci memiliki mata besar, dan sistem optik lebih baik daripada banyak jenis primata, dan mereka juga memiliki sel ganglion pusat / lingkungan standar di retina mereka. Namun, sel-sel standar ini hanya membentuk seperempat dari seluruh populasi sel ganglion retina, diwakili oleh sekitar dua belas tipe fungsional dan anatomi [14,21]. Jelas bahwa masing-masing dari kedua belas jenis sel ini dipenjara oleh evolusi di bawah rangsangan visualnya sendiri. Tetapi pentingnya fakta ini terletak pada fakta bahwa saluran dari sel-sel ganglion non-standar mendominasi aliran visual yang mengalir ke otak kelinci. Informasi ini bukan hal baru: inovasi adalah bukti bahwa retina semua mamalia mentransmisikan berbagai saluran visual pengkodean ke otak [4, 13, 24, 25] dan bahwa keanekaragaman tersebut harus dimasukkan dalam pandangan realistis apa pun dari perangkat visual.
Contoh yang terkenal adalah sel-sel yang secara selektif menanggapi arah pergerakan stimulus. Salah satu dari mereka secara langsung mengirimkan sinyal ke inti batang otak yang terkait dengan gerakan mata [26]. Tetapi tipe lain dari sel-sel ini tidak begitu dikenal, sinyal yang tidak hanya menuju otak tengah, tetapi juga melalui tubuh engkol lateral ke korteks visual primer [19]. Sel-sel seperti itu tidak begitu jarang, tetapi tidak disebutkan di dalam buku teks ketika menggambarkan perangkat penglihatan. Sinyal dari sel-sel ganglion non-standar jelas mencapai korteks visual, tetapi bagaimana cara menggunakannya?
Tipe sel non-standar kedua (diidentifikasi pada kelinci, kucing, dan tikus) adalah detektor tepi lokal. Seperti sel-sel yang selektif responsif terhadap arah gerakan, mereka juga pernah dianggap sebagai sel ganglion langka, tetapi sekarang kita tahu bahwa ini disebabkan oleh kesalahan ketika menggunakan elektroda: mereka mungkin membentuk sekitar 15% dari semua sel ganglion [23]. Sinyal dari sel-sel ini juga mencapai korteks visual primer, tetapi tanggapan mereka jauh melampaui respons visual standar [8,22,23]. Sel-sel ini merespon terbaik terhadap target kecil yang bergerak lambat, seperti predator atau hewan pemangsa, bergerak pada jarak yang jauh. Namun, jika ada banyak objek kecil, seperti pada bidang bertekstur, sel-sel berhenti merespons. Dengan kata lain, sel bereaksi terhadap objek bergerak kecil, tetapi hanya ketika itu sendirian.
Detektor tepi lokal tampaknya merupakan jenis sel ganglion retina yang paling banyak pada kelinci, tetapi tidak termasuk dalam model penglihatan standar. Bisakah sel-sel ini membentuk sistem analisis gerak pada jarak jauh? Bagaimana proses korteks visual memberi sinyal dari sel-sel ini? Bagaimana spesies mamalia lain memandang dunia dengan sel-sel ganglion standar dan detektor tepi lokal - atau bahkan sel dengan kompresi informasi visual lain yang masih harus ditemukan?
Korteks visual lebih pintar daripada yang dikenali buku teks
Bahkan jika kita membuat asumsi bahwa seluruh aliran visual dikodekan hanya oleh sel retina standar, pada saat ini diketahui bahwa pemrosesan dalam korteks visual primer tidak terjadi secara eksklusif sesuai dengan metode yang diusulkan sebelumnya menggunakan neuron "sederhana", "kompleks", "hiper-kompleks", tetapi juga neuron "dengan penghambatan terminal". Diskusi fakta ini telah diberikan banyak ruang dalam literatur khusus [27-30].
Masalahnya dimulai dengan fakta lain yang jelas bahwa hampir semua neuron di korteks serebral adalah non-linear, yang menimbulkan keraguan pada kegunaan skema hierarkis dengan asumsi linear dalam interpretasi fungsi mereka [31,32].
Kedua, gagasan tiga jenis dasar sel - "sederhana", "kompleks" dan "hiper-kompleks" - adalah abstraksi kasar dari beragam jenis sel korteks visual primer, yang diidentifikasi dengan metode anatomi [33,34]. Ketiga, neuron korteks visual bergantung tidak hanya pada aliran visual yang masuk, tetapi juga pada berbagai sinyal kontekstual dari tetangga mereka, sehingga reaksi terhadap rangsangan yang sama yang merupakan bagian dari gambar sehari-hari dan dibuat secara artifisial oleh eksperimen dapat bervariasi [27,35].
Respon dari neuron korteks visual bahkan tidak tetap dalam waktu: pengangkatan sebagian kecil retina, yang mengarah pada munculnya blind spot, memungkinkan neuron yang dibiarkan tanpa bekerja untuk mengubah area tanggung jawab mereka dan memproses informasi visual dari bagian kerja retina yang paling dekat dengan blind spot [36].Dari sudut pandang anatomi, tidak ada yang di atas yang mengejutkan. Hanya hingga 10% dari input neuron individu berasal dari saluran aliran visual [37], sisanya berasal dari neuron tetangga atau dari neuron yang terletak di bagian superior jauh dari korteks serebral, di mana neuron visual masih kurang dipahami dan interpretasi fungsi mereka adalah tugas yang sulit.Dengan demikian, menjadi jelas bahwa pandangan standar perangkat penglihatan terjebak dalam rawa dogma. Langkah apa yang harus diambil untuk keluar dari sana?Langkah 1: tinjau dasar-dasarnya
Berita bagus terbaru adalah bahwa teknik pencitraan sel retina telah mengalami revolusi yang tidak mencolok selama 5 tahun terakhir, sehingga sekarang mudah untuk memvisualisasikannya dalam jumlah besar dan dengan kualitas resolusi yang belum pernah terjadi sebelumnya dalam bentuk gambar tiga dimensi. Masalah menemukan sinapsis dan posisi relatif sel mudah dipecahkan, sekarang mereka dapat diklasifikasikan tidak hanya "oleh mata", tetapi juga menggunakan metode klasifikasi obyektif [15,16]. Tingkat korespondensi pengetikan morfologis dengan pengetikan fisiologis sel retina menunjukkan hubungan langsung (dibahas dalam [24]). Dengan demikian, tipe struktural sel ganglion retina secara akurat menentukan jumlah saluran fungsional dalam aliran visual.Dalam semua sistem visual mamalia yang dipelajari hingga saat ini, data anatomi menunjukkan bahwa jumlah saluran fungsional sekitar dua belas. Fungsi sekitar setengah dari saluran ini dipelajari dengan baik di retina monyet dan kucing. Pada kelinci, proporsi ini sekitar sepertiga, retina tikus dan tikus, meskipun keuntungannya jelas untuk penelitian genetik, sebagian besar tetap belum dijelajahi.Dengan demikian, mungkin berguna untuk kembali, dipersenjatai dengan metode modern, ke bisnis yang belum selesai pada 1970-an [38]. Tugasnya adalah untuk menyelesaikan penentuan fungsi sel ganglion, tahap pertama dari kompresi informasi visual - singkirkan titik-titik buta di awal - dan perbarui tujuan Anda di seluruh sistem visual.Langkah Perbaiki Insentif
Mengapa, setelah lebih dari setengah abad penelitian, adalah pengetahuan fisiologi penglihatan jauh di belakang pengetahuan anatomi, dan apa yang dapat dilakukan untuk memperbaiki situasi? Mengesampingkan masalah selektivitas penelitian menggunakan elektroda, masalah utama tetap menjadi pilihan metode untuk menemukan stimulus yang tepat.Memahami mengapa sel-sel ganglion retina dan neuron dalam korteks visual primer digunakan dalam sistem penglihatan - apa penyesuaian mereka dengan karakteristik lingkungan yang terlihat - jauh lebih sulit daripada yang disarankan oleh para perintis neurosains. Teknik penelitian klasik adalah bahwa para peneliti mendengarkan sinyal amplifikasi dari elektroda yang direndam secara acak di saraf optik atau di korteks serebral sementara secara manual menggerakkan stimulus yang diproyeksikan ke retina untuk mencari area sel tanggung jawab yang disentuh oleh elektroda. Metode cepat dan efektif, tetapi menderita subjektivitas dan kurangnya reproduktifitas. Stimulus kisi sederhana dan analisis sistemik linier sangat efektif untuk sel standar [38,39], tetapi, seperti yang disebutkan sebelumnya,metode ini kurang cocok untuk studi sel non-standar dan neuron non-linear dari korteks visual primer. Upaya baru untuk menentukan serangkaian rangsangan yang benar menarik [40], tetapi sejauh ini belum ada konsensus.Alternatifnya didasarkan pada strategi korelasi terbalik [41-43], di mana stimulus yang dipilih secara acak oleh eksperimen disajikan beberapa kali, dan respon sel digunakan untuk menghitung rata-rata terbalik. Dengan demikian, eksperimen dapat membangun representasi dari rerata rerata yang mengarah ke respons sel paling aktif. Keuntungan besar dari metode ini adalah bahwa hal itu tidak menyiratkan penyetelan spesifik sel terhadap rangsangan tes. Untuk alasan teoretis dan analitis, stimulus uji biasanya dipilih sebagai "acak": papan catur yang berkedip-kedip atau bentuk lain dari kebisingan visual. Strategi ini secara konseptual elegan, tetapi sebagian besar terbatas pada analisis sel dengan organisasi pusat / lingkungan standar,karena sel-sel non-standar (menurut definisi) disesuaikan dengan fitur informasi visual yang sangat nonrandom.Alternatif lain menggunakan pendekatan yang lebih radikal. Strateginya adalah untuk mencari korelasi terbalik dari respon sel terhadap gambar yang diambil pada video dalam lingkungan sehari-hari dan disajikan pada layar monitor [27,44]. Idenya adalah bahwa peneliti mengharapkan sinyal dari sel ketika stimulus penting muncul untuknya. Efisiensi metode semacam itu telah dibuktikan secara eksperimental, tetapi pertanyaannya tetap bagaimana alami gambar dua dimensi harus dipertimbangkan, bagaimana metode ini akan memperhitungkan efek nonlinier dan konteks, dan terlebih lagi bagaimana menafsirkan hasil.Teori visi umum baru?
Perbandingan terbalik dari dunia di sekitar sinyal di otak dapat dilihat sebagai empirisme, diambil secara ekstrem. Tetapi apa yang dapat ditawarkan oleh para ahli teori dari ilmu saraf dibandingkan dengan sesama peneliti? Mereka memiliki peluang untuk berhasil melampaui gaya populer saat ini, dari sudut pandang formal, yang hanya merupakan rumusan ulang hasil eksperimen. Meskipun seperempat abad upaya, sebagian besar pekerjaan yang dilakukan tidak dapat lulus ujian waktu dan tidak dapat dikonfirmasi dalam fakta eksperimental baru. Meskipun formulasi hasil eksperimen yang akurat dan kompak adalah penting, mereka jarang mengarah pada sintesis pengetahuan baru, dan keinginan untuk kenyamanan evaluasi matematis data dapat memaksakan pembatasan yang tidak terlihat pada percobaan. Contoh spesifik diberikan sebelumnya:jika metode eksperimental terbatas pada penyajian parut lebar variabel dalam bentuk stimulus, sel ganglion dengan organisasi tipe "penindasan dengan kontras" hanya muncul sebagai "ledakan" dalam data eksperimental, yang tidak dapat diungkapkan oleh peneliti.Lalu bagaimana para ahli teori dapat membantu? Mereka harus berurusan dengan aspek komputasi yang tidak nyaman dari sistem saraf yang sebenarnya. Sebagai contoh, ahli fisiologi eksperimental tahu betul bahwa sistem sensorik hanya linier ketika eksperimen memaksa mereka menjadi begitu [27,38,45]. Cakupan yang luas dari teori visi diperlukan untuk mengatasi tugas-tugas seperti memahami manfaat pengkodean redundan dan jarang dalam sistem visual, kebutuhan untuk meregangkan jembatan antara studi visi dalam invertebrata dan vertebrata [46,47]. Pertanyaannya adalah apakah memikirkan kembali tahap pertama dalam kompresi informasi visual di retina akan memberikan hasil [46,48-50]. Variasi saluran fungsional dari aliran visual, yang dengan segala keragamannya melayani kita dalam bentuk perasaan yang akrab - visi, sebelumnya ditekankan.Akibatnya, tugas utama akan tetap untuk mencari tahu bagaimana kita hanya "melihat" dan bagaimana secara umum kita bisa memahami ini?Referensi1. Casagrande VA (1994). A third parallel visual pathway to primate area V1. Trends Neurosci. 17, 305–310.
2. Hendry SHC, and Reid RC (2000). The koniocellular pathway in primate vision. Ann. Rev. Neurosci. 23, 127–153.
3. White AJR, Solomon SG, and Martin PR (2001). Spatial properties of koniocellular cells in the lateral geniculate nucleus of the marmoset Callithrix jacchus. J. Physiol. 533, 519–535.
4. Dacey DM, Peterson BB, Robinson FR, and Gamlin PD (2003). Fireworks in the primate retina: in vitro photodynamics reveals diverse LGN-projecting ganglion cell types. Neuron 37, 15–27.
5. Sincich LC, Park KF, Wohlgemuth MJ, and Horton JC (2004). Bypassing V1: a direct geniculate input to area MT. Nat. Neurosci. 7, 1123–1128.
6. Rodieck RW (1967). Receptive fields in the cat retina: a new type. Science 157, 90–92.
7. Stone J. and Hoffmann K.-P. (1972). Very slow-conducting ganglion cells in the cat's retina: a major, new functional type? Brain Res. 43, 610–616.
8. Cleland BG, Levick WR, Morstyn R., and Wagner HG (1976). Lateral geniculate relay of slowly-conducting retinal afferents to cat visual cortex. J. Physiol. 255, 299–320.
9. DeMonasterio FM (1978). Properties of ganglion cells with atypical receptive-field organization in retina of macaques. J. Neurophysiol. 41, 1435–1449.
10. Yukie M., and Iwai E. (1981). Direct projection from the dorsal lateral geniculate nucleus to the prestriate cortex in macaque monkeys. J. Comp. Neurol. 201, 81–97.
11. Hubel DH, and Wiesel TN (1977). Ferrier lecture. Functional architecture of macaque monkey visual cortex. Proc. R. Soc. Lond. Ser. B Biol. Sci. 198, 1–59.
12. Troy JB, Einstein G., Schuurmans RP, Robson JG, and Enroth-Cugell CH (1989). Responses to sinusoidal gratings of two types of very nonlinear retinal ganglion cells of cat. Visual Neurosci. 3, 213–223.
13. Tailby C., Solomon SG, Dhruv NT, Majaj NJ, Sokol SH, and Lennie P. (2007). A new code for contrast in the primate visual pathway. J. Neurosci. 27, 3904–3909.
14. Rockhill RL, Daly FJ, MacNeil MA, Brown SP, and Masland RH (2002). The Diversity of ganglion cells in a mammalian retina. J. Neurosci. 22, 3831–3843.
15. Badea TC, and Nathans J. (2004). Quantitative analysis of neuronal morphologies in the mouse retina visualized by using a genetically directed reporter. J. Comp. Neurol. 480, 331–351.
16. Kong JH, Fish DR, Rockhill RL, and Masland RH (2005). Diversity of ganglion cells in the mouse retina: Unsupervised morphological classification and its limits. J. Comp. Neurol. 489, 293–310.
17. Wassle H. (2004). Parallel processing in the mammalian retina. Nat. Rev. Neurosci. 5, 747–757.
18. Sinha P. (2002). Recognizing complex patterns. Nat. Neurosci. 5 Suppl, 1093–1097.
19. Levick WR, Oyster CW, and Takahashi E. (1969). Rabbit lateral geniculate nucleus: sharpener of directional information. Science 165, 712–714.
20. DeVries SH, and Baylor DA (1997). Mosaic arrangement of ganglion cell receptive fields in rabbit retina. J. Neurophysiol. 78, 2048–2060.
21. Roska B., and Werblin F. (2001). Vertical interactions across ten parallel, stacked representations in the mammalian retina. Nature 410, 583–587.
22. Zeck GM, Xiao Q., and Masland RH (2005). The spatial filtering properties of local edge detectors and brisk-sustained retinal ganglion cells. Eur. J. Neurosci. 22, 2016–2026.
23. van Wyk M., Taylor WR, and Vaney DI (2006). Local edge detectors: a substrate for fine spatial vision at low temporal frequencies in rabbit retina. J. Neurosci. 26, 13250–13263.
24. Masland RH (2001). Neuronal diversity in the retina. Curr. Opin. Neurobiol. 11, 431–436.
25. Dacey DM, Liao HW, Peterson BB, Robinson FR, Smith VC, Pokorny J., Yau KW, and Gamlin PD (2005). Melanopsinexpressing ganglion cells in primate retina signal colour and irradiance and project to the LGN. Nature 433, 749–754.
26. Simpson JI (1984). The accessory optic system. Ann. Rev. Neurosci. 7, 13–41.
27. Carandini M., Demb JB, Mante V., Tolhurst DJ, Dan Y., Olshausen BA, Gallant JL, and Rust NC (2005). Do we know what the early visual system does? J. Neurosci. 25, 10577–10597.
28. Olshausen BA, and Field DJ (2004). Sparse coding of sensory inputs. Curr. Opin Neurobiol. 14, 481–487.
29. Simoncelli EP, and Olshausen BA (2001). Natural image statistics and neural representation. Ann. Rev. Neurosci. 24, 1193–1216.
30. Olshausen BA, and Field DJ (2005). How close are we to understanding v1? Neural. Comput. 17, 1665–1699.
31. Ferster D., and Miller KD (2000). Neural mechanisms of orientation selectivity in the visual cortex. Ann. Rev. Neurosci. 23, 441–471.
32. Anderson JS, Lampl I., Gillespie DC, and Ferster D. (2001). Membrane potential and conductance changes underlying length tuning of cells in cat primary visual cortex. J. Neurosci. 21, 2104–2112.
33. Lund JS, and Yoshioka T. (1991). Local circuit neurons of macaque monkey striate cortex: III. Neurons of laminae 4B, 4A, and 3B. J. Comp. Neurol. 311, 234–258.
34. Cauli B., Audinat E., Lambolez B., Angulo MC, Ropert N., Tsuzuki K., Hestrin S., and Rossier J. (1997). Molecular and physiological diversity of cortical nonpyramidal cells. J. Neurosci. 17, 3894–3906.
35. Zipser K., Lamme VA, and Schiller PH (1996). Contextual modulation in primary visual cortex. J. Neurosci. 16, 7376-7389.
36. Gilbert CD, Sigman M., and Crist RE (2001). The neural basis of perceptual learning. Neuron 31, 681–697.
37. Douglas RJ, and Martin KA (2004). Neuronal circuits of the neocortex. Ann. Rev. Neurosci. 27, 419–451.
38. Troy JB, and Shou T. (2002). The receptive fields of cat retinal ganglion cells in physiological and pathological states: where we are after half a century of research. Prog. Ret. Eye Res. 21, 263–302.
39. Lennie P., and Movshon JA (2005). Coding of color and form in the geniculostriate visual pathway. J. Opt. Soc. Am. 22, 2013–2033.
40. Victor JD (2005). Analyzing receptive fields, classification images and functional images: challenges with opportunities for synergy. Nat. Neurosci. 8, 1651–1656.
41. Srinivasan MV, Jin ZF, Stange G., and Ibbotson MR (1993). 'Vector white noise': a technique for mapping the motion receptive fields of direction-selective visual neurons. Biol. Cybern. 68, 199–207.
42. Meister M., and Berry MJ (1999). The neural code of the retina. Neuron 22, 435–450.
43. Kara P., Reinagel P., and Reid RC (2000). Low response variability in simultaneously recorded retinal, thalamic, and cortical neurons. Neuron 27, 635–646.
44. Ringach DL, Hawken MJ, and Shapley R. (2002). Receptive field structure of neurons in monkey primary visual cortex revealed by stimulation with natural image sequences. J. Vision 2, 12–24.
45. Schwartz O., and Simoncelli EP (2001). Natural signal statistics and sensory gain control. Nat. Neurosci. 4, 819–825.
46. Horridge A. (2000). Seven experiments on pattern vision of the honeybee, with a model. Vision Res. 40, 2589-2603.
47. Srinivasan MV (2006). Honeybee vision: in good shape for shape recognition. Curr. Biol. 16, R58–60.
48. Marr D. (1982). Vision: A Computational Investigation in the Human Representation and Processing of Visual Information (New York: WH Freeman).
49. Barlow HB (1997). The knowledge used in vision and where it comes from. Phil.Trans. Roy. Soc. Lond. B 352, 1141–1147.
50. Riesenhuber M., and Poggio T. (2000). Models of object recognition. Nat. Neurosci. 3, 1199–1204.