
Selama ribuan tahun, umat manusia telah mencari ke surga, ingin menjadi seperti burung. Banyak pemikir hebat, dari Leonardo da Vinci hingga Wright bersaudara, merancang berbagai jenis pesawat, beberapa di antaranya tetap di atas kertas, sementara yang lain menjadi nenek moyang pesawat terbang dan helikopter modern. Sekarang terbang untuk seseorang tidaklah sulit - saya membeli tiket pesawat dan voila. Sebagian besar dari kita bahkan tidak memikirkan bagaimana burung logam besar ini bekerja yang dapat memindahkan kita dari satu benua ke benua lain. Kebenaran antara pesawat modern dan burung asli tidak begitu banyak kesamaan, tetapi dengan robot otonom dikembangkan, situasinya sedikit berbeda. Banyak ilmuwan mencoba membuat robot burung, dan agar inisiatif ini berhasil, perlu dipahami tidak hanya bagaimana burung terbang, tetapi juga bagaimana mendarat. Hari ini kami akan bertemu dengan Anda sebuah studi di mana para lulusan Universitas Stanford mempelajari secara rinci mekanisme penanaman burung nuri bernama Gary dan kerabatnya. Bagaimana burung mendarat, bagaimana mereka memahami bahwa mereka mungkin mendarat atau tidak pada permukaan tertentu, dan apa kinematika anggota tubuh mereka selama pendaratan? Kami akan menemukan jawaban untuk ini dan pertanyaan lain dalam laporan para peneliti. Ayo pergi.
Dasar studi
Apa perbedaan antara pesawat modern dan burung dalam hal pendaratan? Boeing 777, misalnya, membutuhkan landasan pacu dengan panjang setidaknya 2.500 m, helikopter membutuhkan permukaan yang rata (lebih disukai), seperti helikopter. Burung, pada gilirannya, tidak terbatas dalam geometri kanvas tanam dan dapat mendarat di hampir semua permukaan (jika kita menganggap burung sebagai keseluruhan, daripada spesies tunggal). Jika kita berbicara tentang robot otonom, maka kemampuan mereka dalam hal pendaratan sangat terbatas. Beberapa dari mereka dapat berhasil mendarat di permukaan yang dirancang khusus, tetapi ini tidak dapat dibandingkan dengan pelatuk biasa, yang dapat mendarat di permukaan vertikal, miring, horizontal, selama ada cakar yang harus dipegang.
 Ini nyaman bagi saya (semut pelatuk / Melanerpes formicivorus).
Ini nyaman bagi saya (semut pelatuk / Melanerpes formicivorus).Para peneliti mencatat bahwa pendahulu mereka telah melakukan beberapa penelitian yang mengukur dinamika kaki dan sayap burung selama penanaman. Namun, masih belum jelas bagaimana dinamika ini berubah tergantung pada berbagai jenis permukaan yang berfungsi sebagai situs pendaratan. Bagaimanapun, tidak hanya geometri area pendaratan yang penting, tetapi juga tekstur permukaan.
Menarik juga bahwa penelitian sebelumnya tidak dilakukan pada burung hidup, oleh karena itu, dalam praktiknya, tidak mungkin untuk mengevaluasi sepenuhnya kekuatan cengkeraman cakar dan cakar. Itulah sebabnya beberapa pertanyaan tetap tidak jelas: bagaimana tepatnya burung memahami bahwa lokasi pendaratan cocok; apa dinamika anggota badan pada saat menyentuh dan meraih; mengapa burung mendarat di berbagai permukaan adalah masalah sepele, dll.
Dalam pekerjaan yang kami pertimbangkan hari ini, para peneliti memutuskan untuk menguji mekanisme dan dinamika pendaratan burung pohon dengan mengamati beo Lesson (
Forpus coelestis ).
 Budgie Pelajaran.
Budgie Pelajaran.Subjek secara sukarela mendarat di kutub dengan berbagai geometri dan tekstur. Total ada 9 pilihan: 3 "alami", 3 dengan diameter yang berubah dan 3 dengan tekstur yang berubah.
Opsi "Alami" dinamai demikian, karena tempat itu terbuat dari ranting asli (berdiameter 19 mm) dari tiga pohon yang berbeda:
- California Cal - permukaan kasar di berbagai bagian satu cabang;
- ceiba luar biasa - permukaannya relatif halus, rantingnya lunak;
- osmanthus yang harum - permukaannya kasar secara longitudinal, cabang-cabangnya keras.
Untuk percobaan dengan diameter tiang yang dimodifikasi, pin birch biasa dengan diameter berbeda digunakan sebagai bahan:
- 38 mm - diameter terlalu besar untuk jenis nuri ini;
- 19 mm - burung beo dapat melakukan genggaman parsial (jari depan dan belakang tidak bersentuhan);
- 6 mm - burung beo dapat melakukan genggaman penuh (jari depan dan belakang bersentuhan).
Untuk percobaan dengan tekstur yang berbeda, tiang birch dengan diameter 19 mm dibungkus dalam tiga bahan berbeda:
- busa - tekstur lunak dan elastis;
- teflon - tekstur licin;
- Amplas adalah tekstur yang kasar dan kasar.
Parrot sedang menguji semua 9 opsi untuk situs pendaratan.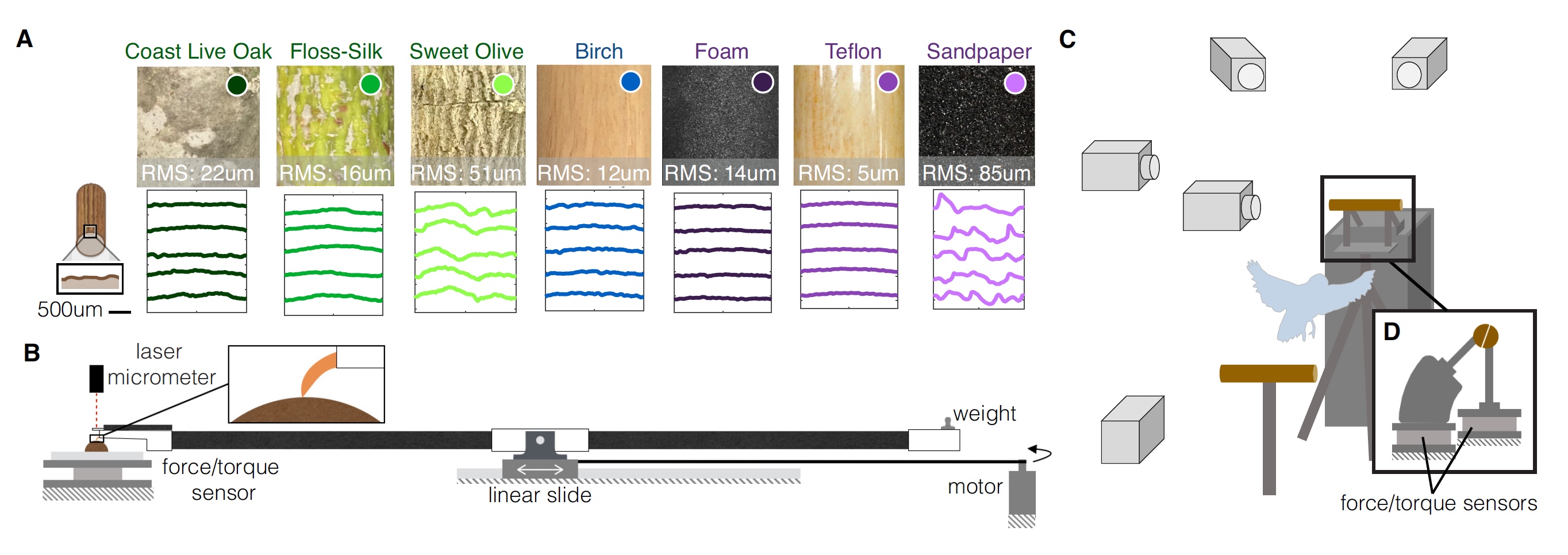 Gambar No. 1
Gambar No. 1Pemindaian cahaya struktural tiga dimensi dari masing-masing varian perforasi dilakukan, yang memungkinkan untuk menggambarkan secara kuantitatif tekstur permukaan masing-masing varian dan membuat profil permukaan (
1A ). Untuk mengevaluasi efek dari fitur-fitur tekstur ini pada interaksi permukaan bertengger dan kaki nuri, para peneliti melakukan uji ketahanan jari dan cakar untuk mengukur kekuatan gesekan untuk setiap permukaan. Tekanan cakar juga dievaluasi untuk mengukur deformasi permukaan (
1B ).
Pada diagram
1C, kita melihat bahwa semua tes dilakukan di bawah pandangan beberapa kamera berkecepatan tinggi, dan kutubnya digergaji menjadi dua. Sensor kekuatan dan momen kekuatan melekat pada masing-masing setengahnya.
Hasil penelitian
Pertama-tama, perhatikan dinamika sayap dan kaki saat mendarat.
Dalam semua pengujian dengan semua varian permukaan pendaratan, subjek menunjukkan dinamika klasik yang sama. Pendaratan dimulai dengan pengereman sayap (fase udara), setelah itu cakar menyerap impuls yang tersisa dalam kontak dengan tenggeran (fase penyerapan). Kontak dibuat dengan kedua kaki dengan penundaan waktu di antara mereka beberapa milidetik, yaitu kaki pertama yang bersentuhan dengan kutub adalah kaki yang disukai:
- Subjek 1 - dalam 100% kasus, kaki kanan;
- Subjek 2 - dalam 83% kasus, kaki kiri;
- Subjek 3 - dalam 85% kasus, kaki kiri.
Setelah kontak, fase pengikatan dimulai, ketika burung beo memperbaiki jari-jarinya di sekitar tempat bertengger dengan lebih andal dan kuat. Ini diikuti oleh tahap yang tidak selalu ditemukan, tetapi seringkali, adalah fase penyesuaian, ketika mereka entah bagaimana mengubah posisi cakar atau jari pada tempat bertengger.
Semua fase pendaratan nuri (sesuai dengan skema pada 2A ). Gambar No. 2
Gambar No. 2Kadang-kadang burung beo, yang tampaknya melebih-lebihkan atau meremehkan kondisi pengujian, sedikit tidak terbang / terbang di atas lokasi pendaratan (
2B ), yang mengarah ke dispersi sudut kemiringan kaki yang lebih besar, di mana mereka membentuk pegangan statis.
Sangat mengherankan bahwa jika ada beberapa variasi dalam indikator kekuatan kaki (
2B ) dan arah (
2C ), nilai rata-rata gaya pendaratan tetap sama untuk berbagai kutub.
Perbedaan dalam indikator yang terlihat selama pendaratan dapat dijelaskan oleh strategi pendaratan burung beo. Ada teori yang menyebutkan burung mengendalikan pendaratannya dengan menilai secara visual waktu kontak dengan permukaan,
τ (
t ). Untuk pendaratan yang sukses, mereka menyesuaikan kecepatan pendekatan mereka untuk mempertahankan konstanta
τ (
t ). Jika sedikit lebih rinci:
τ didefinisikan sebagai jarak ke tempat bertengger dibagi dengan kecepatan perkiraan (
v ).
Jika burung melambat dengan deselerasi konstan (
a ), maka
τ =
s /
v = 0,5
at 2 /
at = 0,5
t , dalam hal ini
τ (
t ) = 0,5.
Jika
τ (
t ) <0,5, penghambatan akan berkurang sampai saat kontak dengan bertengger. Dan jika 0,5 <
τ (
t ) <1, pengereman akan meningkat sampai burung membuat tabrakan terkontrol dengan permukaan pendaratan.
Mengingat kinematika yang dijelaskan di atas, kakaktua mempertahankan nilai
τ (
2E ) yang relatif konstan sesuai dengan nilai tabrakan terkontrol (
2F ). Nilai rata-rata terkecil
τ = 0,80 diamati dalam tes dengan kutub dengan diameter terkecil (6 mm). Dan nilai tertinggi
τ = 0,94 dalam tes dengan kutub yang tertutup busa. Ini menunjukkan bahwa kakatua meningkatkan pengereman saat mendekati permukaan pendaratan lunak.
Kehadiran
τ yang tinggi (mis., Lebih besar dari 0,5) di semua pengujian dengan semua jenis permukaan menunjukkan bahwa burung tidak sepenuhnya mendarat pada saat disentuh.
Setelah berkenalan dengan dinamika pendaratan, kita akan beralih ke kinematika kaki dan cakar teman-teman berbulu kita.
Mengingat keseragaman perilaku selama pendaratan, terlepas dari jenis permukaan, para peneliti menyimpulkan bahwa burung menanggapi perbedaan dalam tekstur dan geometri tiang pendaratan tepat dengan cakar dan cakar mereka, dan bukan sayap mereka. Kedengarannya logis, bukan? Tapi, yang mengejutkan, ini tidak sepenuhnya benar.
Kinematika kaki menunjukkan set stereotip yang sama (sama untuk semua jenis bertengger) dari tahap pendaratan (
3A ,
3B ).
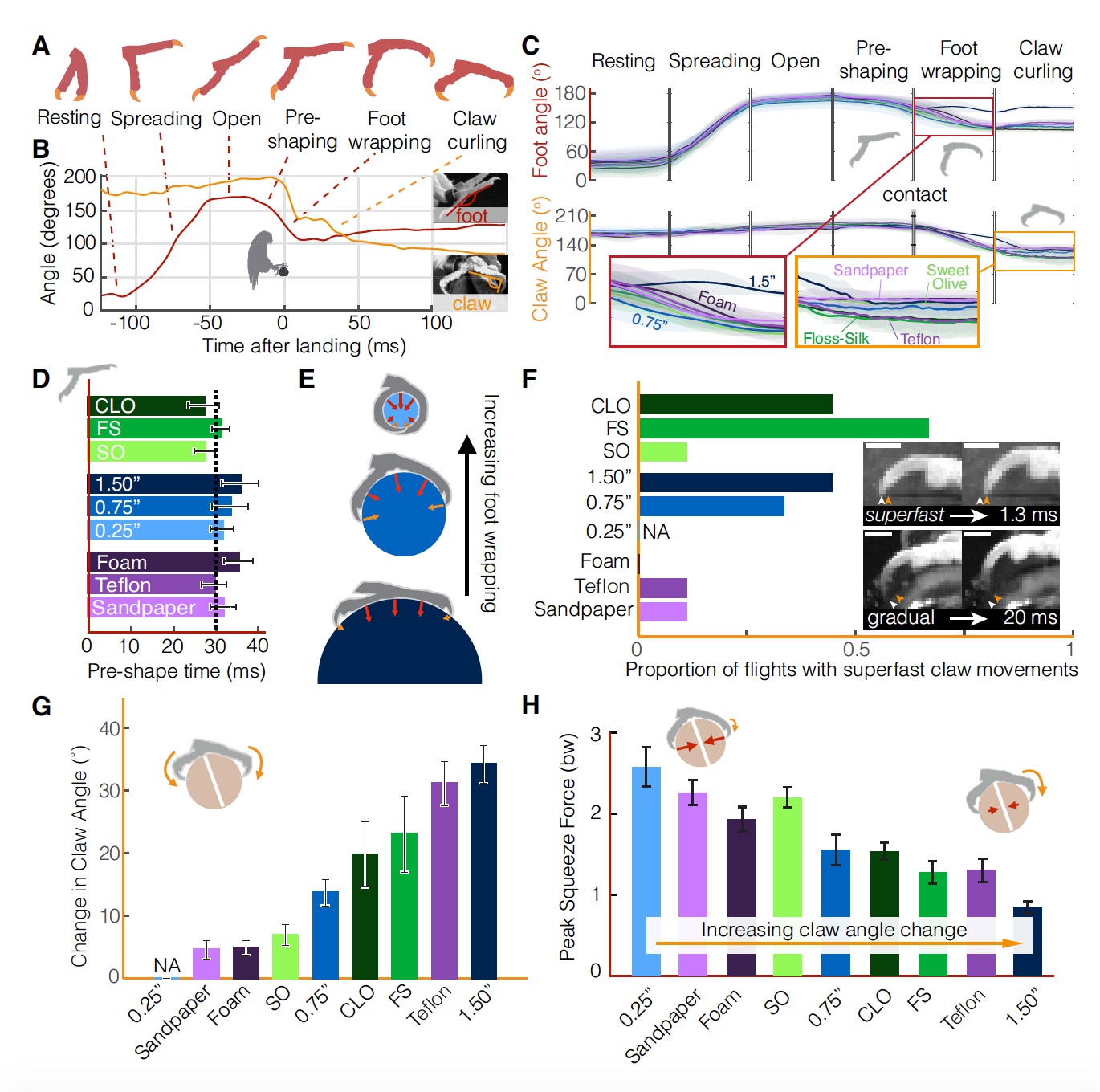 Gambar No. 3
Gambar No. 3Selama penerbangan, kaki dalam keadaan tertutup (fase istirahat), kemudian mereka mulai membuka sekitar 100 ms sebelum menyentuh tempat itu (fase pembukaan berlangsung 40 ± 8 ms), kemudian ada fase "terbuka", ketika dalam 21 ± 7 ms berhenti diungkapkan sepenuhnya.
Segera sebelum kontak, jari-jari mulai mendekati satu sama lain (fase pembentukan awal penangkapan, durasi 31 ± 10 ms). Setelah melakukan kontak dengan permukaan, jari-jari mulai menangkap tenggeran (fase ketebalan, durasi 19 ± 7 ms). Fase terakhir adalah penangkapan cakar yang berlangsung 185 ± 11 ms.
Perbedaan dalam tekstur dan geometri kutub secara eksklusif mempengaruhi kinematika tahapan pendaratan setelah kontak dengan permukaan (
3C ). Sampai kontak, kinematika kaki dan cakar tetap tidak berubah dalam semua tes, bahkan ketika kaki mulai menutup selama fase pra-cengkeraman.
Dengan kata lain, jika burung bereaksi terhadap perbedaan geometri dan tekstur permukaan pendaratan, maka perubahan kinematik akan diamati bahkan selama fase sebelum kontak, yaitu. masih di udara. Sebagai gantinya, kakatua mulai secara berurutan menangkap 30 ms sebelum mendarat (
3D ). Ini mengarah pada asumsi bahwa mereka memiliki solusi yang dapat diprediksi untuk masalah pendaratan di permukaan yang tidak biasa.
Penting bagi burung untuk mempertahankan sudut yang besar di antara cakar sampai saat kontak untuk mencegah kerusakannya. Selanjutnya, tingkat penangkapan ditentukan oleh diameter tiang (
3E ). Pengambilan penuh terjadi dalam kasus kutub dengan diameter 6 mm, mis. burung beo dapat sepenuhnya menggunakan semua jari dan kekuatan cakar untuk menstabilkan.
Penting untuk dicatat bahwa pada saat kontak, cakar dan / atau cakar dapat meluncur dari kutub, karena permukaannya yang halus. Hasilnya, sudut cakar bisa bervariasi. Mempertimbangkan bahwa tahap penjepit cakar dapat bertahan lebih dari 100 ms, pergerakan individual cakar tunggal (
3F ) dapat terjadi dengan sangat cepat (1-2 ms).
Gerakan cakar super cepat mungkin tidak diatur hanya oleh kontraksi otot, para peneliti menyarankan. Juga mengejutkan bahwa di antara vertebrata, gerakan otot tercepat 5-10 kali lebih lambat daripada gerakan cakar super cepat ini (misalnya, otot lengan manusia - 50 ms, otot dada burung kolibri - 8 ms). Dari sini kita dapat menyimpulkan bahwa kecepatan gerakan cakar burung beo tergantung pada pelepasan energi yang disimpan dalam tendon elastis, dan pada inersia cakar yang rendah. Gerakan cakar yang cepat memungkinkan burung beo untuk secara langsung menyesuaikan cengkeramannya tergantung pada geometri dan tekstur permukaan pendaratan.
Adapun tingkat penangkapan cakar, itu tergantung pada diameter tiang dan pada tekstur (
3G ). Pegangan yang paling menonjol diamati dalam kasus diameter 38 mm. Perlu diperjelas bahwa ini hanya tentang menangkap cakar, dan tidak semua kaki sepenuhnya. Artinya, semakin besar diameternya, semakin Anda harus bergantung pada cakar, karena cakarnya tidak akan membantu (
3C ,
3E ). Sudut cakar relatif terhadap permukaan sangat tergantung pada teksturnya. Misalnya, dalam kasus permukaan yang lebih halus (Teflon), cakar "menekuk" lebih banyak (
3H ) daripada dalam kasus permukaan kasar (amplas), karena dalam hal ini mereka dapat dengan mudah menekan permukaan sedikit lebih banyak, sehingga memastikan adhesi yang dapat diandalkan. Secara berlebihan, cakar menekuk lebih keras ketika permukaan lebih sulit dikompres.
 Gambar No. 4
Gambar No. 4Kemudian para peneliti memutuskan untuk memeriksa bagaimana, dari sudut pandang mekanika, indikator kekuatan tekan jari dan tekukan cakar berubah, dan bagaimana mereka mempengaruhi adaptasi burung ke permukaan satu atau lainnya selama pendaratan.
Pengujian dengan tekstur kutub yang berbeda menunjukkan bahwa kedua ujung jari (
4A ) dan cakar (
4C ) menunjukkan
gaya geser yang sama
* pada saat meluncur dan mencengkeram.
Gaya geser * - gaya yang tidak setara mendorong satu bagian tubuh ke satu arah tertentu, dan bagian kedua tubuh di sisi yang berlawanan.
Untuk bantalan kaki, metode yang paling cocok untuk mewakili gaya geser sebagai fungsi gaya normal untuk semua permukaan adalah model gesekan Coulomb (
4B ), yang memungkinkan seseorang untuk memprediksi arah dan besarnya gaya gesekan antara dua benda yang bersentuhan dengan permukaan kering (karenanya model ini juga disebut "model gesekan kering" )
Untuk cakar (
4C ), koefisien gesekan bisa 8 kali lebih tinggi daripada paw pads (
4A ). Perbedaan tersebut dapat disebabkan, antara lain, oleh orientasi cakar relatif terhadap permukaan (
4D ): pada permukaan terlembut, gaya gesekan tetap relatif konstan, tetapi pada permukaan kasar gaya ini juga meningkat dengan meningkatnya sudut cakar. Namun, bahkan pada sudut yang sama, pengukuran menunjukkan bahwa koefisien gaya maksimum untuk cakar masih lebih tinggi daripada untuk bantalan (
4E ).
Untuk mengimbangi sifat stochastic dari adhesi kopling, burung dapat memanfaatkan keseimbangan beban dan pergerakan cakar di permukaan. Distribusi beban antara beberapa cakar secara efektif mempersempit distribusi kekuatan total yang diharapkan dari semua cakar (
4E ). Selain itu, burung dapat memperluas cakar mereka, yang memungkinkan mereka untuk meningkatkan kemungkinan adhesi ke permukaan yang tidak rata, jika ada. Hal ini meningkatkan nilai gaya gesek (
4F ) yang diharapkan, yang menjelaskan tekukan cakar yang lebih jelas pada permukaan yang lebih sulit dikompres dengan kuat (
3F ). Meregangkan cakar memberikan keuntungan dalam hal permukaan di mana ada beberapa penyimpangan, tetapi mereka cukup besar. Namun, pernyataan ini sedikit tidak konsisten dengan pengamatan, karena burung beo tidak terlalu berusaha untuk memperpanjang cakar mereka, dan ini menunjukkan bahwa mereka tidak memilih gundukan terbaik selama penanaman. Karena itu, mereka menekuk cakar mereka sebanyak mungkin hingga kekuatan yang cukup tercapai untuk mempertahankan cengkeraman yang stabil. Jika selama proses ini kekuatan ini tidak tercapai, burung akan mulai menyesuaikan posisi cakar atau lepas landas.
 Gambar No. 5
Gambar No. 5Perbedaan antara koefisien gesekan untuk cakar dan bantalan juga dapat dijelaskan oleh geometri bagian tubuh ini dan geometri permukaan pendaratan yang dengannya mereka bersentuhan. Bantalan memiliki area yang lebih besar dan tekstur lunak, yang memungkinkan Anda mendistribusikan beban di banyak titik kontak, dan ini menyebabkan gaya gesekan yang lebih seragam. Cakar, pada gilirannya, kasar dan memiliki area kecil, oleh karena itu mereka bergantung pada interaksi langsung dengan geometri permukaan, yaitu pada gearing untuk benjolan.
Untuk mengukur efek geometris, para ilmuwan mengkarakterisasi bentuk cakar pada bidang sagital dengan lebar sebagai fungsi ketinggian dari ujung cakar (
5A ). Selain menggunakan penyimpangan permukaan yang ada, cakar juga dapat menghasilkan gaya gesek dari deformasi permukaan.
Untuk mensimulasikan interaksi ini, pengukuran kedalaman penetrasi cakar ke berbagai permukaan (
5B ) dan pengukuran geometri cakar itu sendiri digabungkan. Geometri ujung cakar dimodelkan oleh bola dengan jari-jari 50 μm untuk beban hingga 25% dari berat badan (
5C ). Jadi, ditemukan bahwa, misalnya, kedalaman penetrasi ke dalam tenggeran dari pohon ek California adalah 10 μm, yaitu 0,2% dari panjang busur eksternal cakar (sekitar 5 mm).
Untuk mengevaluasi secara lebih rinci pengaruh ukuran cakar pada traksi, model lain dibuat dengan cakar dengan diameter ujung yang berbeda melekat pada permukaan yang berbeda (
5D ). Seperti yang diharapkan, dengan tidak adanya penetrasi cakar ke permukaan, permukaan yang lebih kasar lebih cocok untuk penangkapan yang sukses (
5E ).
Ketika efek penetrasi permukaan (
5F ) ditambahkan ke model, ditemukan bahwa permukaan yang berguna meningkat. Selain itu, semakin jelas efeknya, semakin kecil sudut yang terbentuk antara sumbu vertikal dan vektor gaya total pada cakar. Jika jari-jari dan massa ujung cakar diskalakan secara isometrik, dan gaya yang diterapkan tergantung pada massa, maka koefisien kedalaman (kedalaman penetrasi dibagi dengan jari-jari ujung cakar) juga akan berskala isometrik.
Dari sini kita dapat membuat kesimpulan logis bahwa penyimpangan kecil pada permukaan ini cukup untuk hewan kecil untuk secara efektif mematuhi permukaan.
 Gambar No. 6
Gambar No. 6Dengan menggabungkan semua model di atas untuk berbagai efek, kekuatan dan hal-hal lain, para peneliti menciptakan model dua dimensi tunggal, yang secara bersamaan memperhitungkan parameter cakar, cakar dan tubuh secara keseluruhan (
6A ).
Totalitas data pada mekanisme kontak kaki dan permukaan pendaratan memungkinkan untuk menerapkan optimasi terbatas untuk menentukan "ruang penangkapan" tiga dimensi burung dan kombinasi kekuatan dan momen kekuatan (
6B ) yang diperlukan. , , : , . , (
6C ).
, . , . , . : , .
, , . , , , . , .
.
Epilog
, , . . , . , , , . , . - -, - — . , , - , . , , , .
Jumat off-top:
( , ), .
Terima kasih atas perhatian Anda, tetap penasaran dan selamat berakhir pekan, semuanya! :)
, . ? ? Dukung kami dengan melakukan pemesanan atau merekomendasikannya kepada teman-teman Anda,
diskon 30% untuk pengguna Habr pada analog unik dari server entry-level yang kami temukan untuk Anda: Seluruh kebenaran tentang VPS (KVM) E5-2650 v4 (6 Cores) 10GB DDR4 240GB SSD 1Gbps dari $ 20 atau bagaimana membagi server? ( RAID1 RAID10, 24 40GB DDR4).
Dell R730xd 2 ? 2 Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 $199 ! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — $99! . c Dell R730xd 5-2650 v4 9000 ?