Automação a partir de DNA e proteínas: o que isso tem em comum com a eletrônica?
(resposta do biólogo ao artigo "Você não escreve animais corretamente" )Existem milhares de genes em todas as células. Nunca é que todos eles trabalhem simultaneamente. As células musculares, hepáticas e cerebrais carregam o mesmo conjunto de genes. Eles são tão diferentes porque genes diferentes trabalham neles. Todas as células podem ativar e desativar genes individuais em resposta a várias influências externas. Ou seja, o sistema que controla a atividade dos genes é um sistema de controle automático. Os cientistas querem entender como essa automação funciona para poder repará-la e quebrá-la. Por exemplo, nas células humanas, existe um sistema de cerca de trezentos genes e proteínas que controlam a divisão celular. Quando se decompõe e a célula começa a se dividir o tempo todo, o câncer ocorre.Um diagrama simplificado da rede de genes que regula a divisão das células humanasComo é a automação intracelular?
No "ferro" é geralmente usado impulsos elétricos. Os sinais na biologia são geralmente químicos, ou seja, mudanças na concentração de qualquer substância. Existem sinais elétricos no sistema nervoso, mas este é apenas um dispositivo para transmissão rápida de longa distância, semelhante à fibra óptica em tecnologia. A transmissão de um impulso nervoso de célula para célula ocorre em forma química, memória de longo prazo e a complexa integração de sinais em um neurônio também é química.Onde houver sinais, haverá pistas - os sinais estão fora do lugar e o ruído é lixo aleatório grudando nos sinais. Em eletrônica, a chegada de sinais no endereço é fornecida por fios e isolamento. A diafonia ocorre devido ao acoplamento indutivo capacitivo e indutivo entre os condutores. Em uma célula bacteriana, seu conteúdo é mais ou menos uniformemente misturado e qualquer sinal químico (e existem milhares deles) está disponível em qualquer um de seus pontos. As células de animais e plantas são divididas em compartimentos com composição química diferente, mas os tipos desses compartimentos são inferiores a uma dúzia e cada um contém centenas e milhares de substâncias diferentes.A chegada do sinal no lugar certo na célula é devida ao reconhecimento molecular do princípio do "bloqueio de teclas". Uma molécula de proteína pode reconhecer outra proteína, uma proteína pode reconhecer uma sequência de DNA específica, uma proteína pode reconhecer pequenas moléculas como açúcares. Esse reconhecimento nem sempre é perfeitamente preciso; portanto, moléculas com formato semelhante a um parceiro de reconhecimento regular criarão dicas. Além da interferência, existem ruídos na eletrônica. Eles surgem devido ao movimento térmico dos átomos, que introduz uma fração do caos no movimento dos elétrons. Em uma célula, o ruído químico também está associado ao movimento térmico. Quando o sinal é fraco, por exemplo, dez pedaços de moléculas desse tipo por toda a célula, essas moléculas serão espalhadas pela célula aleatoriamente, e não pelo fato uniforme. Como resultado, sua concentração em algum canto da célula irá flutuar aleatoriamente, e isso será ruído.O que exatamente é regulado em uma célula viva?
Uma gaiola pode ser considerada como uma planta que produz todo o equipamento necessário e pode montar um segundo da mesma planta. O principal equipamento são os ribossomos - máquinas CNC que coletam todas as proteínas. A estrutura da proteína coletada é registrada em um RNA de fita (mRNA). O ribossomo se move ao longo do mRNA e, de acordo com suas instruções, coleta uma cadeia de uma nova proteína a partir de unidades individuais - aminoácidos. A cadeia de proteínas emergente do ribossomo é dobrada em uma bola compacta de proteína madura, que começa a funcionar. As proteínas podem acelerar as reações químicas (enzimas), bem como envolver-se no transporte de substâncias, sinalização, defesa e muitas outras coisas.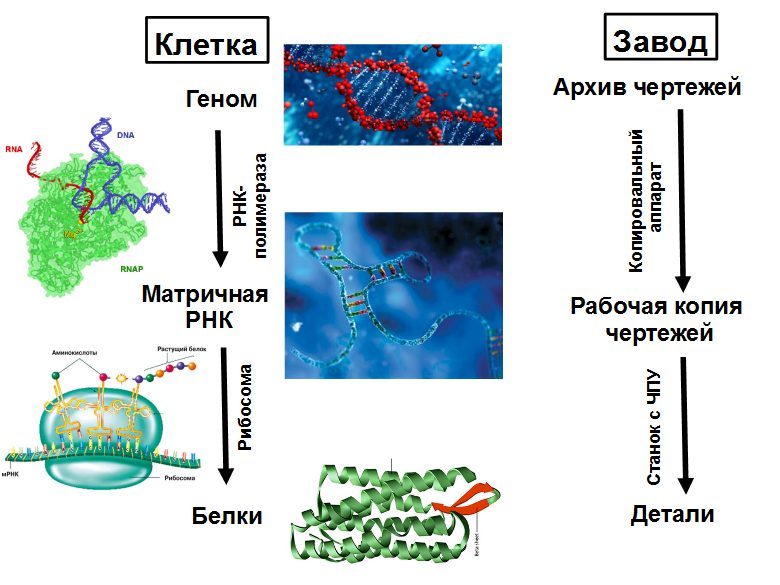
Elementos lógicos no DNA e proteínas
Talvez o exemplo mais estudado de mudança de gene seja o operon lactose de Escherichia coli. O francês Jacob e Mono por sua pesquisa receberam o Prêmio Nobel em 1965. E. coli, como o próprio nome indica, vive no intestino. Pode ser o intestino de uma grande variedade de animais, de abelhas a humanos. Lá, ela tem que comer o que o proprietário recebeu, para que ela possa comer muitos nutrientes diferentes.E. coli sob um microscópio e em uma placa de Petri AE. coli pode crescer, por exemplo, em um meio nutritivo composto apenas por sais de glicose e minerais e produzir todos os aminoácidos e vitaminas de que precisa. Em vez de glicose, pode haver outros açúcares (frutose, açúcar de malte - maltose, açúcar de leite - lactose e uma dúzia de outros), além de polissacarídeos, como amido.Para absorver cada um desses açúcares, você precisa de suas próprias enzimas. A produção dessas enzimas deve ser iniciada apenas quando o açúcar correspondente estiver no ambiente, caso contrário, os materiais e a energia para a produção dessas enzimas serão desperdiçados. Ou seja, os genes dessas enzimas de digestão de açúcar só devem ser ativados quando esse açúcar estiver presente. Para assimilar cada açúcar, via de regra, são necessárias várias enzimas, e não uma. Seus genes estão localizados nas proximidades da cadeia de DNA e sua atividade é controlada por uma região reguladora comum do DNA no início do primeiro gene. Esse grupo de genes colaborativos e controlados de forma síncrona é chamado de "operon".Esquema do operon lactose Ooperon lactose consiste em três genes. O primeiro (LacZ) codifica uma proteína de transporte que bombeia a lactose para a célula, e os outros dois (LacI e LacA) codificam enzimas, cujo trabalho conjunto transforma a lactose em "açúcar padrão" - glicose. No início do operon lactose, existe um local de ligação à RNA polimerase (promotor) e um local de ligação à proteína reguladora (operador).A atividade do operon lactose é controlada por dois sinais. O primeiro sinal, obviamente, é a concentração de lactose. Se não houver lactose, as enzimas de sua assimilação não são necessárias. O segundo sinal é um pouco mais complicado. Em um experimento, Jacob e Mono cultivaram E. coli em um meio contendo lactose e glicose. Sob tais condições, o bastão primeiro consome glicose e seu operon lactose é desligado. Quando a glicose termina, o crescimento de bactérias pára por 15 a 20 minutos e depois continua devido à lactose. Durante uma pausa, o operon lactose é ativado. Ou seja, a segunda entrada reguladora desliga o operon lactose quando a célula possui mais açúcares disponíveis do que a lactose (a captação de glicose não requer enzimas adicionais).Experiências posteriores mostraram que o operon lactose não responde à glicose propriamente dita. Em vez disso, o nível de fome (mais precisamente, a "qualidade" dos açúcares disponíveis) dentro da célula de E. coli é codificado por uma substância sinalizadora especial. É chamado de "adenosina monofosfato cíclico" ou cAMP. Enquanto o bastão cresce com glicose, praticamente não existe cAMP na célula. Se não houver glicose, mas houver maltose ou amido (um pouco mais difícil de assimilar as substâncias), um pequeno cAMP é produzido. Se não houver maltose, mas houver frutose ou lactose, o nível de cAMP na célula será maior. Se não houver açúcar, mas houver ácido lático - ainda mais alto e, finalmente, se não houver nada saboroso e você tiver que comer glicerina, o nível de AMPc será o mais alto. Assim, dois sinais controlam o operon da lactose: concentração de lactose e concentração de cAMP.Como esses sinais afetam a atividade dos genes? Um gene ativo é aquele em que a RNA polimerase produz muitos RNAs mensageiros. A RNA polimerase sempre começa a trabalhar nesse operon aterrissando na mesma região de DNA (é chamada de "promotor"). Onde é o lugar para a lactose e o cAMP? A RNA polimerase não pode reconhecer todas as centenas de sinais que regulam a atividade de milhares de genes. Para isso, existem proteínas reguladoras especiais (elas são chamadas de "fatores de transcrição"). Duas proteínas reguladoras estão envolvidas no gerenciamento do operon lactose, uma para cada sinal de entrada: um repressor de lactose e um ativador catabólico.Repressor de lactose (roxo) no DNA Umrepressor de lactose é uma proteína que pode se ligar à lactose ou a uma sequência específica de DNA. Existem duas dessas seqüências no genoma de Escherichia coli, ambas no início do operon lactose, em ambos os lados do promotor. As moléculas repressoras de lactose aderem firmemente umas às outras em grupos de quatro moléculas de proteína. A ligação de um desses quatro ao DNA torce a fita de DNA em um circuito fechado no qual o promotor termina. A RNA polimerase não pode se apoiar em um promotor tão distorcido e o gene se desliga. Se a lactose aparecer na célula, ela se liga ao repressor da lactose e cai do DNA, liberando o promotor da RNA polimerase - o operon começa a funcionar. É assim que a entrada reguladora do operon pela concentração de lactose é organizada.Um ativador catabólico no DNA Umativador catabólico responde de maneira semelhante à concentração de AMPc. É verdade que existem várias diferenças nos detalhes:- o ativador catabólico fica no DNA, exatamente ao lado do promotor e não se sobrepõe. Pelo contrário, aumenta a ligação da RNA polimerase ao promotor e aumenta a atividade do operon.- um ativador catabólico liga cAMP e DNA. Sem cAMP, ele não pode se ligar ao DNA.- é necessário um ativador catabólico para a atividade completa do operon lactose, mas mesmo sem ele, o operon continuará funcionando por cerca de 5% da atividade total. O repressor de lactose regula o operon na faixa de 0 a 100%.- As moléculas do ativador catabólico são conectadas em pares, não em quatro. No próximo post, veremos a que essa diferença leva.Ou seja, podemos escrever uma tabela verdade para o operon lactose:O operon lactose funciona quase como um elemento lógico AND. Apenas 5% da atividade no estado de "lactose +, cAMP -" cai ligeiramente da imagem ideal.Continua com gráficos e matan: https://geektimes.ru/post/283686/ Source: https://habr.com/ru/post/pt399233/
All Articles