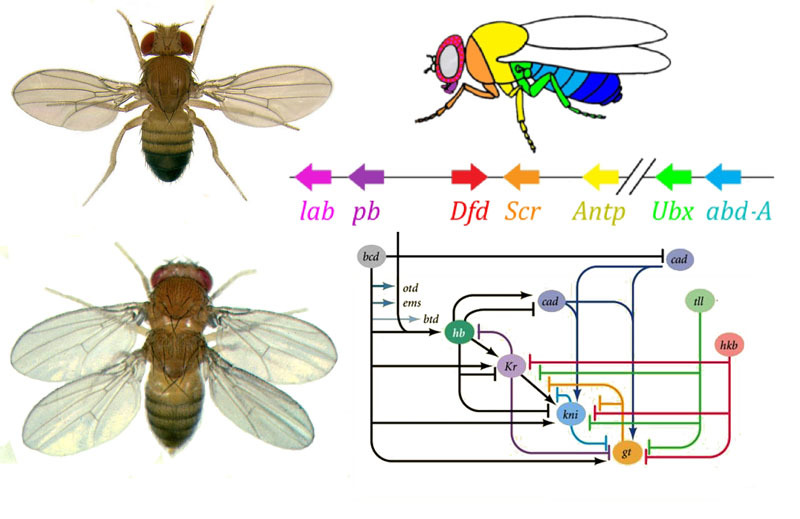
 Como usar um ovo para transformar um ovo em larva e depois em mosca adulta? Uma mosca, como outros insetos, consiste em segmentos. Os segmentos são combinados em três grupos - cabeça, tórax e abdômen. Um segmento pode ter um par de membros. Existem três segmentos no peito, cada um com um par de pernas. O segundo segmento torácico também tem asas, e o terceiro - o besouro do solo (pequenas asas da cauda, quase invisíveis sem uma lupa). O abdômen é composto por oito segmentos; não há membros neles. Inicialmente, havia seis segmentos na cabeça, mas durante a evolução dos insetos os limites entre eles foram completamente apagados. Seus membros são antenas com as quais a mosca cheira e sente e três pares de membros da boca. Para um inseto ou uma barata, seriam mandíbulas, maxilas e lábio inferior, enquanto que para uma mosca todas elas se fundem na probóscide.Para que todos os órgãos estejam no lugar, cada célula deve saber onde está localizada no embrião. Portanto, nos estágios iniciais do desenvolvimento embrionário, quando ainda não existem órgãos e todas as células parecem iguais, uma “grade coordenada” bioquímica aparece nela com os eixos ântero-posterior, dorso-abdominal e esquerdo-direito. Mutações dos genes que formam a grade de coordenadas podem alterar fácil e rapidamente a aparência do animal - mover órgãos para outro local, alterar seu número e assim por diante. Portanto, esses genes são importantes para a evolução dos animais e são muito ativamente estudados. A rede de genes para marcar o embrião de uma mosca de Drosophila, o brinquedo favorito dos geneticistas, é talvez a mais estudada entre as redes de genes de animais.O algoritmo para marcar um ovo ao longo do eixo anteroposterior é basicamente simples:1) determine qual extremidade do ovo será a cabeça, qual cauda2) divida o embrião ao longo do eixo anteroposterior em 17 segmentos3) divida cada segmento nas partes frontal, média e traseira4) marque a cabeça (6 segmentos), tórax (3 segmentos) e abdômen (8 segmentos)5) marque as diferenças individuais dos segmentos Arede de genes que lida com isso possui um dispositivo modular. Cada módulo executa sua própria etapa do algoritmo geral. A mosca Drosophila acabou sendo um objeto muito conveniente para o estudo de redes de desenvolvimento de genes, porque possui deformidades hereditárias muito brilhantes e reveladoras. Por exemplo, antennapedia de mutação em que as antenas são transformados em pernas ou
Como usar um ovo para transformar um ovo em larva e depois em mosca adulta? Uma mosca, como outros insetos, consiste em segmentos. Os segmentos são combinados em três grupos - cabeça, tórax e abdômen. Um segmento pode ter um par de membros. Existem três segmentos no peito, cada um com um par de pernas. O segundo segmento torácico também tem asas, e o terceiro - o besouro do solo (pequenas asas da cauda, quase invisíveis sem uma lupa). O abdômen é composto por oito segmentos; não há membros neles. Inicialmente, havia seis segmentos na cabeça, mas durante a evolução dos insetos os limites entre eles foram completamente apagados. Seus membros são antenas com as quais a mosca cheira e sente e três pares de membros da boca. Para um inseto ou uma barata, seriam mandíbulas, maxilas e lábio inferior, enquanto que para uma mosca todas elas se fundem na probóscide.Para que todos os órgãos estejam no lugar, cada célula deve saber onde está localizada no embrião. Portanto, nos estágios iniciais do desenvolvimento embrionário, quando ainda não existem órgãos e todas as células parecem iguais, uma “grade coordenada” bioquímica aparece nela com os eixos ântero-posterior, dorso-abdominal e esquerdo-direito. Mutações dos genes que formam a grade de coordenadas podem alterar fácil e rapidamente a aparência do animal - mover órgãos para outro local, alterar seu número e assim por diante. Portanto, esses genes são importantes para a evolução dos animais e são muito ativamente estudados. A rede de genes para marcar o embrião de uma mosca de Drosophila, o brinquedo favorito dos geneticistas, é talvez a mais estudada entre as redes de genes de animais.O algoritmo para marcar um ovo ao longo do eixo anteroposterior é basicamente simples:1) determine qual extremidade do ovo será a cabeça, qual cauda2) divida o embrião ao longo do eixo anteroposterior em 17 segmentos3) divida cada segmento nas partes frontal, média e traseira4) marque a cabeça (6 segmentos), tórax (3 segmentos) e abdômen (8 segmentos)5) marque as diferenças individuais dos segmentos Arede de genes que lida com isso possui um dispositivo modular. Cada módulo executa sua própria etapa do algoritmo geral. A mosca Drosophila acabou sendo um objeto muito conveniente para o estudo de redes de desenvolvimento de genes, porque possui deformidades hereditárias muito brilhantes e reveladoras. Por exemplo, antennapedia de mutação em que as antenas são transformados em pernas ou VROTMNENOGIproboscipedia, quando parte da probóscide se transforma em pernas. A mutação no bórax dá uma mosca com quatro asas, que além das asas usuais no segundo segmento torácico, possui um segundo par de asas no terceiro segmento torácico.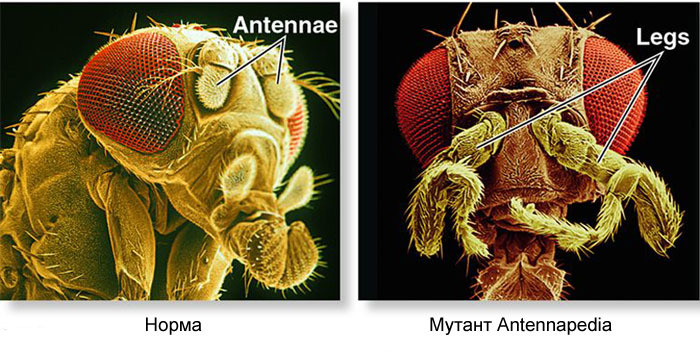
 Tais mutações, nas quais uma parte do corpo se torna muito semelhante a outra, são chamadas de "homeose". Começando com esses mutantes, a genética finalmente decifrou toda a rede de genes que controlam o desenvolvimento da mosca. Este trabalho ganhou o Prêmio Nobel em 1995.Tudo começa com os genes da polaridade do ovo - bicoide e nanos. Eles trabalham mesmo antes da fertilização do ovo, enquanto ele amadurece no ovário da mosca da mãe. As células auxiliares, bombeando nutrientes para o óvulo, estão localizadas em um lado do óvulo, de modo que o óvulo tem duas extremidades diferentes desde o início. O RNA do bicoide da matriz se acumula na extremidade anterior do ovo, onde está ligado e não pode nadar para longe. O nanos mRNA também é fixado na extremidade posterior. Após a fertilização, as proteínas começam a ser sintetizadas usando esses mRNAs. Suas moléculas se espalham por todo o ovo. Como a síntese das proteínas bicoide e nanos ocorre em uma extremidade do ovo e decai em todos os lugares, um gradiente de proteína bicoide diminui, diminuindo da extremidade anterior do embrião para a posterior e o gradiente nano da extremidade posterior para a anterior.
Tais mutações, nas quais uma parte do corpo se torna muito semelhante a outra, são chamadas de "homeose". Começando com esses mutantes, a genética finalmente decifrou toda a rede de genes que controlam o desenvolvimento da mosca. Este trabalho ganhou o Prêmio Nobel em 1995.Tudo começa com os genes da polaridade do ovo - bicoide e nanos. Eles trabalham mesmo antes da fertilização do ovo, enquanto ele amadurece no ovário da mosca da mãe. As células auxiliares, bombeando nutrientes para o óvulo, estão localizadas em um lado do óvulo, de modo que o óvulo tem duas extremidades diferentes desde o início. O RNA do bicoide da matriz se acumula na extremidade anterior do ovo, onde está ligado e não pode nadar para longe. O nanos mRNA também é fixado na extremidade posterior. Após a fertilização, as proteínas começam a ser sintetizadas usando esses mRNAs. Suas moléculas se espalham por todo o ovo. Como a síntese das proteínas bicoide e nanos ocorre em uma extremidade do ovo e decai em todos os lugares, um gradiente de proteína bicoide diminui, diminuindo da extremidade anterior do embrião para a posterior e o gradiente nano da extremidade posterior para a anterior.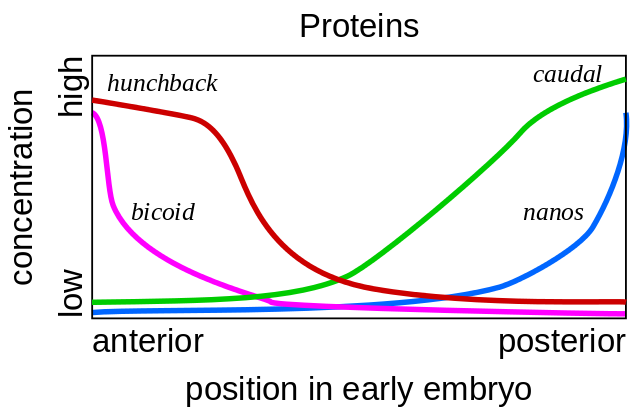 Se um mRNA de bicoide for introduzido em um local diferente no ovo com uma micropipeta, uma segunda cabeça crescerá lá. Se o mRNA do bicoide é desativado artificialmente, um embrião é obtido sem a cabeça e com dois jumentos.Além disso, as proteínas bicoide e nanos incluem os genes do próprio embrião, isto é, um novo mRNA começa a ser lido a partir deles). Os genes gap são os primeiros a começar a trabalhar, a partir da lacuna inglesa, a lacuna. Os dois primeiros genes desse grupo - corcunda e caudal - formam gradientes da cabeça à cauda (corcunda) e da cauda à cabeça (caudal). Mas seus gradientes são mais suaves que os dos bicoides e nanos.
Se um mRNA de bicoide for introduzido em um local diferente no ovo com uma micropipeta, uma segunda cabeça crescerá lá. Se o mRNA do bicoide é desativado artificialmente, um embrião é obtido sem a cabeça e com dois jumentos.Além disso, as proteínas bicoide e nanos incluem os genes do próprio embrião, isto é, um novo mRNA começa a ser lido a partir deles). Os genes gap são os primeiros a começar a trabalhar, a partir da lacuna inglesa, a lacuna. Os dois primeiros genes desse grupo - corcunda e caudal - formam gradientes da cabeça à cauda (corcunda) e da cauda à cabeça (caudal). Mas seus gradientes são mais suaves que os dos bicoides e nanos.Por que os genes são tão estranhamente chamados?. . , hunchback — «», tailless — «», kruppel - — «», bithorax – « ». Sonic hedgehog , .
Os quatro genes restantes do grupo gap formam, cada um, uma ou duas bandas largas (mais de um segmento) com limites bastante claros. Eles são incorporados por produtos proteicos de genes superiores (bicoide, nanos, corcunda e caudal). Um desligamento completo de um dos genes desse grupo produz um embrião encurtado, que carece de vários segmentos, naqueles em que esse gene normalmente trabalhava.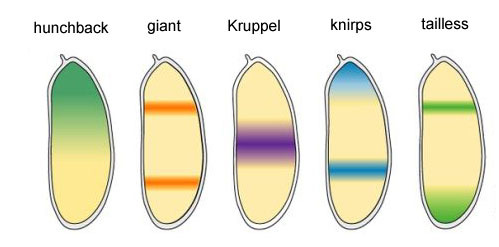 Áreas de atividade dos genes de bloqueio de gapPor que acontece que quatro genes de bloqueio de gap (kruppel, gigante, knirps e sem cauda) dividem o embrião em faixas, em cada um dos quatro genes que funciona?Em primeiro lugar, todos suprimem a atividade um do outro. Em uma única célula, uma condição seria estável, na qual um gene desse grupo trabalha ativamente e suprime todos os outros (gatilho). Mas o ovo da mosca em desenvolvimento é mais complexo. Nele, neste momento, milhares de núcleos celulares individuais flutuam no líquido intracelular total (citoplasma). O tamanho do ovo é grande o suficiente para que a taxa de decaimento das proteínas reguladoras seja comparável à sua taxa de difusão na outra extremidade do ovo. Com uma combinação de difusão e supressão mútua, outro estado estável aparece quando o ovo é dividido em várias bandas, em cada uma das quais um gene do grupo trabalha. Quanto mais lenta a difusão de proteínas ou mais rápida a quebra, menores serão essas faixas ou pontos.A participação de tais sistemas (chamados de reação-difusão) na marcação do corpo dos animais foi prevista por Alan Turing (sim, que inventou a "máquina de Turing") em 1952. Naquela época, pouco se sabia sobre genes e proteínas, mas as equações que descrevem estruturas listradas e malhadas foram deduzidas corretamente por Turing.Aqui ou aqui, você pode brincar com um modelo simples de difusão de reação de duas substâncias e sentir como ele pode se comportar.
Áreas de atividade dos genes de bloqueio de gapPor que acontece que quatro genes de bloqueio de gap (kruppel, gigante, knirps e sem cauda) dividem o embrião em faixas, em cada um dos quatro genes que funciona?Em primeiro lugar, todos suprimem a atividade um do outro. Em uma única célula, uma condição seria estável, na qual um gene desse grupo trabalha ativamente e suprime todos os outros (gatilho). Mas o ovo da mosca em desenvolvimento é mais complexo. Nele, neste momento, milhares de núcleos celulares individuais flutuam no líquido intracelular total (citoplasma). O tamanho do ovo é grande o suficiente para que a taxa de decaimento das proteínas reguladoras seja comparável à sua taxa de difusão na outra extremidade do ovo. Com uma combinação de difusão e supressão mútua, outro estado estável aparece quando o ovo é dividido em várias bandas, em cada uma das quais um gene do grupo trabalha. Quanto mais lenta a difusão de proteínas ou mais rápida a quebra, menores serão essas faixas ou pontos.A participação de tais sistemas (chamados de reação-difusão) na marcação do corpo dos animais foi prevista por Alan Turing (sim, que inventou a "máquina de Turing") em 1952. Naquela época, pouco se sabia sobre genes e proteínas, mas as equações que descrevem estruturas listradas e malhadas foram deduzidas corretamente por Turing.Aqui ou aqui, você pode brincar com um modelo simples de difusão de reação de duas substâncias e sentir como ele pode se comportar.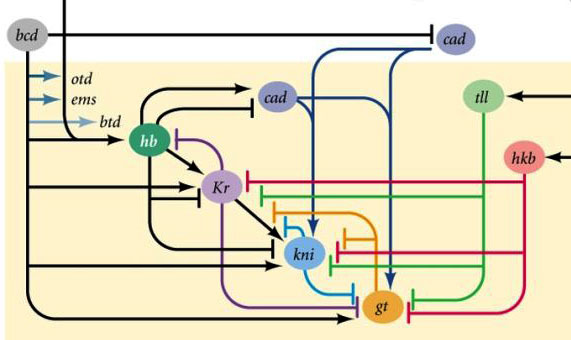 Ligações entre genes de gap e genes de polaridade de ovo. Bcd - bicoide, hb - corcunda, cad - caudal, Kr - Kruppel, kni - knop, gt - gigante, sem cauda.A supressão dos genes um do outro por si só marcará o embrião em algumas faixas, mas a localização das faixas de atividade dos quatro genes em relação um ao outro e nas extremidades do embrião será muito variável. Para que as tiras estejam sempre no lugar (por exemplo, para joelheiras - uma faixa na cabeça e a outra na metade traseira entre o kruppel e tiras gigantes), você precisa de um segundo sinal de controle - gradientes de bicoide e outras proteínas de polaridade do ovo. As funções de entrada dos genes gap para esses reguladores têm máximos em diferentes níveis do sinal de entrada (derivamos essa função de entrada no final do segundo artigo ). Portanto, os genes de gap são ativados pelo nível de proteínas bicoide, corcunda e caudal, e suas zonas de atividade estão naturalmente localizadas da cabeça à cauda do embrião.Depois que os genes do gap fizeram uma marcação aproximada do embrião em 7 bandas (eles não coincidem com os segmentos ou com a cabeça-peito-abdômen), o próximo módulo é ativado - regra dos pares ("regras das regras dos pares"). Ele já divide o embrião em segmentos, que devem ter 17. O módulo de regra de pares consiste em cinco genes, e todos eles, como você deve ter adivinhado, se suprimem. Cada um desses genes funciona em todos os segmentos pares ou ímpares - as proteínas codificadas por eles se decompõem mais rapidamente do que as proteínas do grupo gap, e as tiras, respectivamente, são mais estreitas. Desligar parte dos genes desse grupo produz um embrião abreviado, no qual todos os segmentos pares ou ímpares desaparecem.
Ligações entre genes de gap e genes de polaridade de ovo. Bcd - bicoide, hb - corcunda, cad - caudal, Kr - Kruppel, kni - knop, gt - gigante, sem cauda.A supressão dos genes um do outro por si só marcará o embrião em algumas faixas, mas a localização das faixas de atividade dos quatro genes em relação um ao outro e nas extremidades do embrião será muito variável. Para que as tiras estejam sempre no lugar (por exemplo, para joelheiras - uma faixa na cabeça e a outra na metade traseira entre o kruppel e tiras gigantes), você precisa de um segundo sinal de controle - gradientes de bicoide e outras proteínas de polaridade do ovo. As funções de entrada dos genes gap para esses reguladores têm máximos em diferentes níveis do sinal de entrada (derivamos essa função de entrada no final do segundo artigo ). Portanto, os genes de gap são ativados pelo nível de proteínas bicoide, corcunda e caudal, e suas zonas de atividade estão naturalmente localizadas da cabeça à cauda do embrião.Depois que os genes do gap fizeram uma marcação aproximada do embrião em 7 bandas (eles não coincidem com os segmentos ou com a cabeça-peito-abdômen), o próximo módulo é ativado - regra dos pares ("regras das regras dos pares"). Ele já divide o embrião em segmentos, que devem ter 17. O módulo de regra de pares consiste em cinco genes, e todos eles, como você deve ter adivinhado, se suprimem. Cada um desses genes funciona em todos os segmentos pares ou ímpares - as proteínas codificadas por eles se decompõem mais rapidamente do que as proteínas do grupo gap, e as tiras, respectivamente, são mais estreitas. Desligar parte dos genes desse grupo produz um embrião abreviado, no qual todos os segmentos pares ou ímpares desaparecem.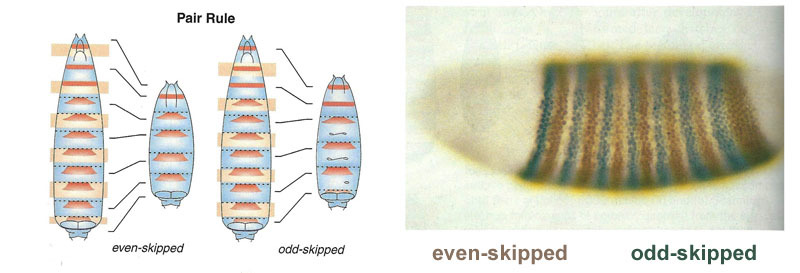 Zonas de atividade de genes do grupo de regras de pares. A foto à direita tem 14 faixas, não 17, porque os outros três sistemas de marcação funcionam nos três primeiros segmentos da cabeça.O próprio módulo de regra de pares, sem a participação de gap e bicoide, pode marcar o embrião em tiras estreitas, mas o número dessas tiras seria instável: os segmentos podem ser 12, 18 e até 20. Normalmente, os genes das regras de pares são difíceis de controlar lacuna de genes e a primeira marcação aproximada (para 7 zonas) ajuda a fazer a marcação final com precisão e confiabilidade (para 14 + 3 segmentos).O próximo ponto do algoritmo é marcar a frente, o meio e o verso de cada segmento. Isso é feito por genes de polaridade de segmento. Eles também se suprimem e são ativados por módulos superiores da rede de genes (regra de pares e gap). Somente os parâmetros de difusão e deterioração de seus produtos são selecionados para que as tiras sejam muito finas - metade de um segmento. Suprimi-los um com o outro é mais fraco e mais organizado do que com genes de regras de pares. Portanto, suas tiras de atividade se sobrepõem parcialmente e em cada segmento é obtida uma “linha” frequente de tiras diferentemente sobrepostas.A desativação dos genes de polaridade do segmento geralmente não leva ao encurtamento do embrião e à perda de suas partes (como é típico para lacunas e regras de pares), mas a efeitos mais sutis. Normalmente, nesse caso, metade de cada segmento no qual esse gene deveria funcionar se transforma em uma imagem espelhada da outra metade. Isso é claramente visível, porque a larva da mosca tem fileiras de pequenos espinhos direcionados para trás - para a conveniência de rastejar. Mutantes nos genes de polaridade do segmento têm fileiras de espinhos direcionados para frente.Finalmente, é a vez dos genes mais famosos e descobertos dessa rede - o módulo hox. O trabalho deles dá individualidade aos segmentos, de modo que as antenas e probóscide cresçam nos segmentos da cabeça e as pernas e asas nos segmentos torácicos. Os genes hox, diferentemente dos módulos anteriores, não inibem a atividade um do outro e podem trabalhar juntos no mesmo segmento embrionário. Uma propriedade estranha dos genes hox que ainda não foi explicada: eles estão localizados no cromossomo em um grupo restrito e na mesma ordem em que trabalham no corpo do embrião.
Zonas de atividade de genes do grupo de regras de pares. A foto à direita tem 14 faixas, não 17, porque os outros três sistemas de marcação funcionam nos três primeiros segmentos da cabeça.O próprio módulo de regra de pares, sem a participação de gap e bicoide, pode marcar o embrião em tiras estreitas, mas o número dessas tiras seria instável: os segmentos podem ser 12, 18 e até 20. Normalmente, os genes das regras de pares são difíceis de controlar lacuna de genes e a primeira marcação aproximada (para 7 zonas) ajuda a fazer a marcação final com precisão e confiabilidade (para 14 + 3 segmentos).O próximo ponto do algoritmo é marcar a frente, o meio e o verso de cada segmento. Isso é feito por genes de polaridade de segmento. Eles também se suprimem e são ativados por módulos superiores da rede de genes (regra de pares e gap). Somente os parâmetros de difusão e deterioração de seus produtos são selecionados para que as tiras sejam muito finas - metade de um segmento. Suprimi-los um com o outro é mais fraco e mais organizado do que com genes de regras de pares. Portanto, suas tiras de atividade se sobrepõem parcialmente e em cada segmento é obtida uma “linha” frequente de tiras diferentemente sobrepostas.A desativação dos genes de polaridade do segmento geralmente não leva ao encurtamento do embrião e à perda de suas partes (como é típico para lacunas e regras de pares), mas a efeitos mais sutis. Normalmente, nesse caso, metade de cada segmento no qual esse gene deveria funcionar se transforma em uma imagem espelhada da outra metade. Isso é claramente visível, porque a larva da mosca tem fileiras de pequenos espinhos direcionados para trás - para a conveniência de rastejar. Mutantes nos genes de polaridade do segmento têm fileiras de espinhos direcionados para frente.Finalmente, é a vez dos genes mais famosos e descobertos dessa rede - o módulo hox. O trabalho deles dá individualidade aos segmentos, de modo que as antenas e probóscide cresçam nos segmentos da cabeça e as pernas e asas nos segmentos torácicos. Os genes hox, diferentemente dos módulos anteriores, não inibem a atividade um do outro e podem trabalhar juntos no mesmo segmento embrionário. Uma propriedade estranha dos genes hox que ainda não foi explicada: eles estão localizados no cromossomo em um grupo restrito e na mesma ordem em que trabalham no corpo do embrião.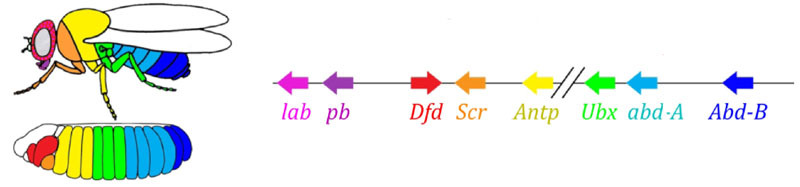 A marcação do embrião é baseada no princípio "quanto mais distante do peito, mais genes hox e mais ativo": a atividade do gene A abdominal-A começa com 2 segmentos abdominais, aumenta até 4 e continua no nível máximo em todos os segmentos subsequentes; O gene abdominal-B trabalha do 5º ao 8º segmento do abdome, aumentando a atividade em cada segmento subsequente. O gene ultrabitórax está incluído na meia-força no terceiro segmento torácico e com capacidade total em todo o abdômen. Quando desativada nos mutantes do bórax, as células do 3 segmento torácico não sabem que são o 3 segmento torácico, elas têm o mesmo conjunto de genes hox que no 2 torácico (ou seja, apenas antennapedia) e o desenvolvimento do 3 segmento torácico prossegue programa 2 no peito - com asas em vez de besouros no chão e poderosos músculos voadores.Se você desativar todos os genes hox em um segmento separado, ele ficará feio, mas acima de tudo parecerá um peito, com pernas e sem asas. Isso ocorre porque os ancestrais distantes dos insetos eram como trilobitas ou milípedes. Eles tinham muitos segmentos idênticos com as pernas. Posteriormente, seu corpo encurtou, a parte de trás perdeu as pernas, tornando-se o abdômen, e vários segmentos da frente uniram-se à cabeça e as pernas tornaram-se partes do aparelho oral (mandíbulas, maxilas e lábio inferior).
A marcação do embrião é baseada no princípio "quanto mais distante do peito, mais genes hox e mais ativo": a atividade do gene A abdominal-A começa com 2 segmentos abdominais, aumenta até 4 e continua no nível máximo em todos os segmentos subsequentes; O gene abdominal-B trabalha do 5º ao 8º segmento do abdome, aumentando a atividade em cada segmento subsequente. O gene ultrabitórax está incluído na meia-força no terceiro segmento torácico e com capacidade total em todo o abdômen. Quando desativada nos mutantes do bórax, as células do 3 segmento torácico não sabem que são o 3 segmento torácico, elas têm o mesmo conjunto de genes hox que no 2 torácico (ou seja, apenas antennapedia) e o desenvolvimento do 3 segmento torácico prossegue programa 2 no peito - com asas em vez de besouros no chão e poderosos músculos voadores.Se você desativar todos os genes hox em um segmento separado, ele ficará feio, mas acima de tudo parecerá um peito, com pernas e sem asas. Isso ocorre porque os ancestrais distantes dos insetos eram como trilobitas ou milípedes. Eles tinham muitos segmentos idênticos com as pernas. Posteriormente, seu corpo encurtou, a parte de trás perdeu as pernas, tornando-se o abdômen, e vários segmentos da frente uniram-se à cabeça e as pernas tornaram-se partes do aparelho oral (mandíbulas, maxilas e lábio inferior).Modelos matemáticos da rede genética para marcação de embriões
No caso do operon lactose e dos loops de ligação direta em Escherichia coli, é possível construir modelos matemáticos bastante simples com um número moderado de parâmetros que prevêem o comportamento dos genes para quaisquer sinais de entrada. Para um operon lactose, basta:- quatro constantes de ligação (repressor e ativador com DNA, lactose com repressor, cAMP com ativador)- quatro parâmetros estequiométricos (4 lactose por 1 repressor, 2 ativador cAMP 1, 1 repressor e ativador por operon)- dois parâmetros de regulação (o efeito do repressor e ativador na probabilidade de aterrissagem da RNA polimerase)- o nível básico de atividade do operon (quando não há repressor ou ativador)- atraso na transcriçãoTotal de 12 parâmetros.Vamos agora olhar para as regiões reguladoras dos genes Hox, digamos. No diagrama abaixo, um dos blocos reguladores do gene B abdominal. As barras coloridas indicam locais reguladores de ligação às proteínas. Todo esse bloco tem um tamanho de cerca de 1700 nucleotídeos - 10 vezes mais que a região reguladora do operon lactose. Possui 39 locais de plantio para 7 proteínas reguladoras diferentes de todos os blocos superiores dessa rede de genes. E o abdominal-B tem quatro desses blocos regulatórios. Eles ativam sequencialmente do quinto ao oitavo segmento do abdômen e criam um aumento gradual do nível de proteína B abdominal nesses segmentos. Ou seja, apenas para o modelo matemático desse gene, em boa medida, são necessárias mais de 100 constantes de ligação, o mesmo número de parâmetros de regulação e um número desconhecido de parâmetros para descrever a interação entre diferentes entradas - na pior das hipóteses, até 1.500 interações pareadas entre todos os locais de desembarque de proteínas em um unidade reguladora.Ou seja, o modelo completo dessa rede de genes certamente terá mais de mil parâmetros. E este é o principal problema: não há tantos sinais de saída que possamos medir no experimento - de fato, apenas os níveis de cada uma das 25 proteínas e sua distribuição ao longo do comprimento do embrião. Portanto, ninguém tentou simular essa rede com a mesma granularidade do operon lactose.Você pode tentar simplificar a tarefa e descrever o efeito de cada proteína reguladora no gene com apenas três parâmetros (parâmetro de ligação, estequiometria e regulação). Você pode selecionar esses parâmetros (por exemplo, usando cadeias de Markov) para que o modelo reproduza com precisão as faixas reais da atividade do gene. Nos dois primeiros blocos da rede (genes de polaridade dos ovos e genes de diferença), esses modelos são feitos ( por exemplo ).O problema é que diferentes partidas do algoritmo de seleção fornecem parâmetros de modelo diferentes - mas todos funcionam corretamente e reproduzem a atividade dos genes em um verdadeiro broto de mosca! Esses modelos têm características comuns (por exemplo, todos os genes de gap suprimem-se e são ativados pelas proteínas-mãe), mas não há menos diferenças do que semelhanças.E como diferentes tentativas de modelagem dão resultados diferentes, é improvável que pelo menos uma delas coincida com a realidade. Pior, esses modelos reproduzem a operação normal, mas perdem muito em prever os resultados das mutações.Na próxima parte, falaremos sobre por que esses problemas com o estudo das redes de genes são inevitáveis em biologia (e a rede de marcação de embriões de Drosophila é uma das mais simples de estudar!), O que pode ser feito sobre isso e o que engenheiros devem aprender com os biólogos.
As barras coloridas indicam locais reguladores de ligação às proteínas. Todo esse bloco tem um tamanho de cerca de 1700 nucleotídeos - 10 vezes mais que a região reguladora do operon lactose. Possui 39 locais de plantio para 7 proteínas reguladoras diferentes de todos os blocos superiores dessa rede de genes. E o abdominal-B tem quatro desses blocos regulatórios. Eles ativam sequencialmente do quinto ao oitavo segmento do abdômen e criam um aumento gradual do nível de proteína B abdominal nesses segmentos. Ou seja, apenas para o modelo matemático desse gene, em boa medida, são necessárias mais de 100 constantes de ligação, o mesmo número de parâmetros de regulação e um número desconhecido de parâmetros para descrever a interação entre diferentes entradas - na pior das hipóteses, até 1.500 interações pareadas entre todos os locais de desembarque de proteínas em um unidade reguladora.Ou seja, o modelo completo dessa rede de genes certamente terá mais de mil parâmetros. E este é o principal problema: não há tantos sinais de saída que possamos medir no experimento - de fato, apenas os níveis de cada uma das 25 proteínas e sua distribuição ao longo do comprimento do embrião. Portanto, ninguém tentou simular essa rede com a mesma granularidade do operon lactose.Você pode tentar simplificar a tarefa e descrever o efeito de cada proteína reguladora no gene com apenas três parâmetros (parâmetro de ligação, estequiometria e regulação). Você pode selecionar esses parâmetros (por exemplo, usando cadeias de Markov) para que o modelo reproduza com precisão as faixas reais da atividade do gene. Nos dois primeiros blocos da rede (genes de polaridade dos ovos e genes de diferença), esses modelos são feitos ( por exemplo ).O problema é que diferentes partidas do algoritmo de seleção fornecem parâmetros de modelo diferentes - mas todos funcionam corretamente e reproduzem a atividade dos genes em um verdadeiro broto de mosca! Esses modelos têm características comuns (por exemplo, todos os genes de gap suprimem-se e são ativados pelas proteínas-mãe), mas não há menos diferenças do que semelhanças.E como diferentes tentativas de modelagem dão resultados diferentes, é improvável que pelo menos uma delas coincida com a realidade. Pior, esses modelos reproduzem a operação normal, mas perdem muito em prever os resultados das mutações.Na próxima parte, falaremos sobre por que esses problemas com o estudo das redes de genes são inevitáveis em biologia (e a rede de marcação de embriões de Drosophila é uma das mais simples de estudar!), O que pode ser feito sobre isso e o que engenheiros devem aprender com os biólogos.