Na minha publicação anterior , duas questões foram consideradas: os fundamentos da biologia molecular e os métodos para criar as construções de que precisamos para genes baseados em vetores plasmídicos. Agora você precisa descobrir como introduzir um vetor plasmídeo em uma célula bacteriana, ou seja, para fazer uma "transformação". Ao mesmo tempo, aprenderemos algo sobre a estrutura da membrana bacteriana e como ela pode ser superada, bem como por que algumas bactérias são chamadas competentes, como tudo isso está associado a toxinas bacterianas e resistência bacteriana a antibióticos, e muitos outros fatos interessantes.
Portanto, temos um vetor plasmídico em nossas mãos e, como existe uma cepa bacteriana adequada para esse vetor, resta introduzir o vetor na bactéria.
Em um
artigo anterior , o
trabalho de Frederick Griffith foi mencionado, o que demonstra claramente o fato de que o DNA pode penetrar espontaneamente nas células bacterianas. A capacidade de uma célula bacteriana de absorver uma molécula de DNA ambiental é chamada de "competência" (que gera muitas piadas sobre o fato de que mesmo unicelulares in vitro são competentes, mas seu colega de laboratório não é). Se a absorção não ocorrer sob a influência de algumas condições criadas artificialmente, estamos falando de "competência natural". Como ocorre um transporte de DNA semelhante dentro de uma bactéria?
1) A estrutura da membrana celular de bactérias
Para começar, consideramos brevemente a estrutura da membrana de bactérias gram-positivas e gram-negativas.
A casca de uma bactéria gram-negativa consiste em duas bicamadas lipídicas: a membrana externa e a membrana interna (a interna também é chamada de plasma). Entre eles está o espaço periplásmico preenchido com proteínas e peptidoglicano, desempenhando funções importantes.
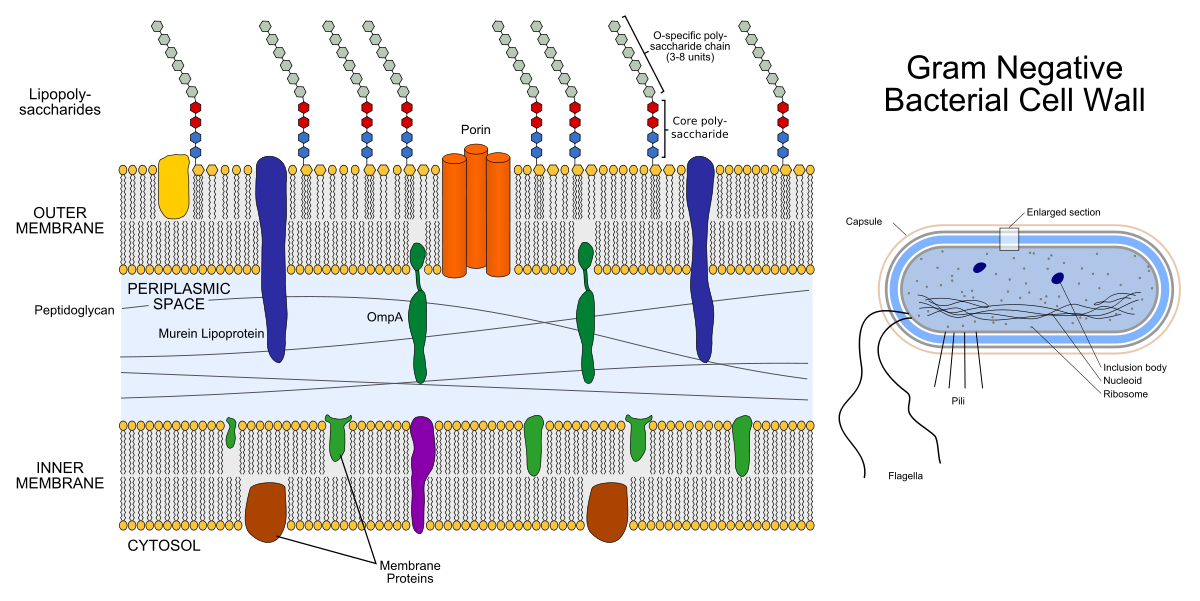 À esquerda está a estrutura da membrana da bactéria gram-negativa. Os aparelhos coloridos dentro das membranas são proteínas da membrana e as cadeias que se projetam são lipopolissacarídeos. À direita está a estrutura da própria bactéria gram-negativa.
À esquerda está a estrutura da membrana da bactéria gram-negativa. Os aparelhos coloridos dentro das membranas são proteínas da membrana e as cadeias que se projetam são lipopolissacarídeos. À direita está a estrutura da própria bactéria gram-negativa.O peptidoglicano é um heteropolímero que consiste em N-acetilglucosamina reticulada covalentemente (NAG) e ácido N-acetilmurâmico (NAM). Acontece que a cadeia ....- NAG-NAM-NAG-NAM-NAG- .... "Hetero" na palavra "heteropolímero" significa que o polímero não consiste nos mesmos elementos (como o polietileno), mas em dois diferentes. A enzima sintetizadora do peptidoglicano, a transpeptidase, é um alvo dos antibióticos β-lactâmicos (antibióticos da penicilina e cefalosporinas): eles inibem sua atividade, como resultado do processo de divisão bacteriana, não há peptidoglicano suficiente para as duas células filhas. A camada peptidoglicana também é um alvo para a enzima lisozima: destrói a ligação covalente entre NAG e NAM. Na medicina, a lisozima é usada como anti-séptico, por exemplo, é um componente ativo de alguns medicamentos para o tratamento de dores de garganta, e na indústria de alimentos pode ser usada como conservante (suplemento alimentar E1105 - essa é a lisozima).

O envelope das bactérias gram-positivas consiste apenas em uma membrana interna (plasma), um espaço periplásmico muito fino e uma camada externa de peptidoglicano mais espessa do que as bactérias gram-negativas.
Tais obstáculos que impedem qualquer composto no citoplasma bacteriano são intransponíveis para tudo, exceto a água, bem como pequenas moléculas não polares e hidrofóbicas (oxigênio molecular e nitrogênio, dióxido de carbono e outros). A penetração desses compostos através da casca é chamada de "transporte passivo" ("difusão simples"), sempre ocorre ao longo do gradiente de concentração. Moléculas não polares são capazes de passar livremente através das membranas, porque o volume interno das membranas também é não polar (dentro da membrana são compostas de resíduos hidrofóbicos de ácidos graxos) e compostos não polares se dissolvem bem em solventes não polares (álcool, acetona, ácidos graxos).
Uma imagem cômica na qual a hidrofobicidade (hidrofobia) dos resíduos de ácidos graxos dentro de uma membrana de duas camadas é reproduzida.Por sua vez, grandes moléculas polares (aminoácidos, monossacarídeos, nucleotídeos), partículas carregadas (íons) e macromoléculas (DNA, proteínas) não conseguem penetrar através da membrana celular assim, pois são bem solúveis apenas em solventes polares (por exemplo, na água) . Então, como as moléculas de DNA entram na célula bacteriana?
2) Mecanismos de transporte induzido de compostos para as células do meio ambiente
Em primeiro lugar, temos em nossas mãos o fato óbvio de que a bactéria não pode viver apenas de água, oxigênio e dióxido de carbono; para a vida, ela precisa de muitos outros compostos. Portanto, existem maneiras de entregar esses compostos na célula. Tais métodos são "difusão facilitada" e "transporte ativo".
2.1) Difusão de luz
Um exemplo de difusão facilitada (embora esse caso em particular seja prejudicial às bactérias) é o transporte de íons potássio através da membrana de uma célula bacteriana em combinação com a valinomicina, um antibiótico sintetizado por alguns fungos. A valinomicina livre não pode penetrar na célula, pois os grupos polares são expostos externamente. O íon de potássio também é um "caminho fechado".
 Uma escultura representando a estrutura tridimensional de um complexo de valinomicina e íon potássio (o íon é representado como uma bola no centro). Foi instalado em frente à entrada do M.M. Shemyakin e Yu. A. Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences (Moscow), no qual foram estabelecidos o mecanismo da atividade antibacteriana da valinomicina e a estrutura tridimensional de seu complexo com íons de potássio.
Uma escultura representando a estrutura tridimensional de um complexo de valinomicina e íon potássio (o íon é representado como uma bola no centro). Foi instalado em frente à entrada do M.M. Shemyakin e Yu. A. Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences (Moscow), no qual foram estabelecidos o mecanismo da atividade antibacteriana da valinomicina e a estrutura tridimensional de seu complexo com íons de potássio.Porém, após a formação de um complexo de valinomicina com um íon potássio, ele altera sua estrutura tridimensional para que toda a superfície externa se torne hidrofóbica (não polar) e todos os grupos hidrofílicos sejam ocultos no interior. Nesse caso, o íon potássio fica oculto dentro da molécula de valinomicina; portanto, o complexo valinomicina + íon potássio pode penetrar através da membrana. O complexo pode se decompor em valinomicina e íons livres a qualquer momento, tanto dentro como fora da célula, mas a difusão geralmente facilitada de íons leva à equalização de sua concentração nos espaços intracelular e extracelular, e a célula controla o transporte de íons por uma razão: sua alta concentração fatal. É sobre isso que se baseia o efeito antibacteriano da valinomicina.
Representação esquemática do antibiótico valinomicina fúngico complexado com íon potássio.Outro exemplo são os poros não controlados, que são grandes proteínas transmembranares que permitem que determinadas substâncias passem livremente pela membrana de acordo com o gradiente. Um exemplo desse poro transmembranar é a toxina α do
Staphylococcus aureus (outro nome para a toxina é α-hemolisina. Consiste nas partes "hemo" e "lisina". "Hemo" significa glóbulos vermelhos; "lisina" significa da palavra "lise", ou seja, "morte celular")
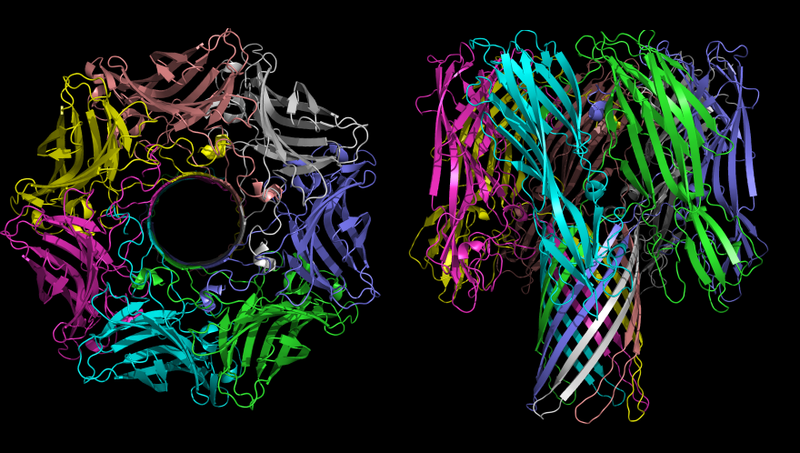 A estrutura tridimensional do oligômero α-hemolisina - a toxina da bactéria Staphylococcus aureus. Na imagem à esquerda, a hora é claramente visível.
A estrutura tridimensional do oligômero α-hemolisina - a toxina da bactéria Staphylococcus aureus. Na imagem à esquerda, a hora é claramente visível.A principal função da α-hemolisina no processo patogênico é a criação de canais não controlados na membrana celular, que, dependendo das condições ambientais, podem transmitir íons monovalentes, íons cálcio ou ATP. Como mencionado acima, o desequilíbrio iônico é extremamente tóxico e o transporte de ATP
da célula é prejudicial, uma vez que o ATP é um elo indispensável em quase todos os processos químicos de qualquer célula.
2.2) Transporte ativo
Como pode ser visto nos exemplos acima, a difusão facilitada não requer energia. Pelo contrário, transporte ativo é a absorção ou liberação de algo por uma célula com o gasto de energia para cada ato de transferência. Pode ser o transporte de grandes moléculas polares ou íons, e o transporte de íons pode ser realizado ao longo do gradiente de concentração e contra ele. Obviamente, o transporte ativo é um processo controlado no qual todo um conjunto de proteínas intracelulares e transmembranares está envolvido. Assim, primeiro a célula precisa gastar recursos na síntese de todas essas proteínas e, depois, novamente gastar energia no uso desse sistema de transporte.
Um exemplo de transporte ativo é o sistema de transporte dependente de energia de compostos hidrofóbicos e anfifílicos de uma célula para o meio ambiente (efluxo).
Sistema de transporte ativo de um antibiótico de uma célula bacteriana para o exterior, consistindo em três proteínas que formam um único complexo transmembranar. A figura reflete o fato de que a proteína transportadora de efluxo (violeta) pode "capturar" moléculas de droga diretamente no citoplasma e na membrana.Esse sistema é de particular interesse, pois causa a multirresistência de algumas bactérias aos antibióticos: a célula se livra deles antes que eles possam prejudicá-lo seriamente. Além disso, mais da metade dos casos de resistência a
múltiplas drogas de
Pseudomonas aeruginosa Pseduomonas aeruginosa está associada ao transporte ativo de antibióticos. Mais importante, já foram descobertos sistemas ativos de efluxo de "novos" antibióticos usados contra bactérias resistentes a medicamentos antibacterianos comumente usados. Por exemplo, o medicamento
Linezolid usado no caso de ineficácia dos antibióticos clássicos é revertido pelo sistema de efluxo baseado nas proteínas AcrAB e TolC. Outro fato interessante é que uma bactéria inteligente não desperdiça energia na síntese em larga escala de proteínas do sistema de efluxo naquelas situações em que não são necessárias. Mas assim que substâncias nocivas para a célula aparecem, a síntese é rapidamente ativada e a bactéria começa a resistir ativamente. A propósito, o efluxo não é o único mecanismo para proteger bactérias de antibióticos, também escreverei um artigo sobre outros métodos.
A estrutura do linezolida é um antibiótico usado contra cepas resistentes de bactérias gram-positivas.3) Competência de células bacterianas
Assim, examinamos os mecanismos de transporte transmembranar de compostos que não podem penetrar nessa barreira sem assistência externa. Sob condições naturais, o transporte de DNA para a bactéria é realizado apenas através do transporte ativo, devido à cooperação de todo um espectro de proteínas intracelulares e transmembranares. A capacidade não induzida de uma célula de absorver o DNA de fora é chamada de "competência natural".
3.1) Competência natural das células bacterianas
A hipótese mais fundamentada do surgimento da competência natural das bactérias é a suposição de que esse mecanismo permita que as bactérias sobrevivam em condições extremas para elas (condições extremas para uma célula são chamadas de "estresse"). O fato é que o estresse geralmente leva a danos no DNA "cromossômico" de uma bactéria e, se uma célula deseja sobreviver, precisa, de alguma forma, "reparar" seu "cromossomo" (esse "reparo" é chamado de "reparo"). Além disso, sabe-se que, se uma sequência de DNA é introduzida na célula que é semelhante a alguma parte do próprio DNA da célula, é altamente provável que essa parte "nova" seja incorporada ao seu genoma. O que acontece sob condições estressantes? Algumas células morrem, tornando-se doadoras de DNA para os irmãos que ainda lutam pela vida. De fato, dentro da mesma cultura bacteriana, é muito provável que os genomas dos representantes sejam muito semelhantes, de modo que as bactérias sobreviventes possam usar o DNA dos parentes mortos para reparar seu genoma.
Além disso, sob condições confortáveis na cultura de células, apenas uma fração delas possui competência natural, e a proporção de células competentes de seu número total é a maior então; a cultura está em uma certa fase de crescimento - a fase
logarítmica .
 Uma parcela típica de bactérias em cultura ao longo do tempo. A fase logarítmica é a fase de crescimento, seguida por um platô e a fase de extinção, na qual as células já carecem de nutrientes e toxinas se acumulam.
Uma parcela típica de bactérias em cultura ao longo do tempo. A fase logarítmica é a fase de crescimento, seguida por um platô e a fase de extinção, na qual as células já carecem de nutrientes e toxinas se acumulam.Obviamente, o pesquisador gostaria de tornar o processo de transferência do vetor plasmídeo para a célula o mais eficiente possível: quanto mais eficiente o processo de transformação, menos vetor plasmídeo é necessário para garantir bons resultados. Portanto, em condições de laboratório, eles geralmente não dependem da própria célula, mas criam células com competência artificial.
3.2) Competência artificial de células bacterianas
Existem dois métodos principais para criar a competência artificial de bactérias: tratamento de uma cultura celular com íons divalentes, seguido de aquecimento a curto prazo e a chamada eletroporação.
3.2.1) Tratamento da cultura celular com íons divalentes, seguido de aquecimento a curto prazo.
Como regra, no laboratório há um refrigerador com uma temperatura de cerca de -80 graus Celsius (às vezes eles são chamados de "kelvinators" em homenagem a uma das empresas de manufatura)), entupidos com tubos de ensaio com células na fase
logarítmica . Isso é feito para não perder tempo criando-os cada vez que forem necessários (e leva muito tempo, não funcionará mais rápido do que em um dia). E para que durante o armazenamento as células não morram devido ao congelamento, crioprotetores são adicionados ao meio: glicerol, propileno glicol ou dimetilsulfóxido.
Refrigerador de laboratório na visão do autor da imagem.Quando um pesquisador precisa transformar uma cepa específica, ele remove as células do Kelvinator e coloca o tubo no gelo. Então, cátions divalentes e o vetor plasmídeo de que precisamos são adicionados às células na fase
logarítmica do crescimento. O tubo de ensaio não deve deixar o gelo. Um doador típico desses cátions é o cloreto de cálcio (CaCl2); nesse caso, nosso cátion divalente é o Ca2 +. Não há explicação inequívoca de como o Ca2 + ajuda o DNA a penetrar na bactéria:
- Algumas fontes afirmam que altas concentrações de Ca2 + causam lacunas na membrana celular e danos ao DNA, e anteriormente descobrimos que o dano ao DNA é um sinal para que a célula se torne competente;
- De acordo com outras fontes, esses íons servem como uma "ponte" que liga o vetor plasmídeo à célula. O fato é que os íons carregados positivamente se ligam simultaneamente a grupos de polissacarídeos carregados negativamente na membrana externa das bactérias e a grupos carregados negativamente de DNA plasmídico. Se não houvesse íons no meio, o DNA seria repelido da célula, e isso é completamente inútil para o pesquisador.
É possível que ambas as hipóteses sejam verdadeiras.
O suposto mecanismo de participação dos íons Ca2 + na transformação. Primeiro, os íons se ligam a grupos de DNA com carga negativa (círculos amarelos) e polissacarídeos ancorados na membrana (círculos amarelos). Então, devido ao íon, forma-se um tipo de ponte: "polissacarídeo (-)" - "íon Ca (2+)" - "DNA (-)", que fixa o DNA na superfície da célula.Após a incubação bacteriana a uma temperatura próxima de zero, é produzido o chamado choque térmico - aquecimento a curto prazo da cultura a 42 graus. Normalmente, a duração do tratamento térmico é de 30 a 45 segundos, após o qual o tubo de ensaio com bactérias retorna ao banho de gelo (com um choque térmico mais longo, as células podem morrer). As possíveis razões para a eficácia do choque térmico são os mesmos mecanismos: a alta temperatura pode causar danos ao DNA e às proteínas (isso induz a competência) e também pode criar "buracos" na membrana celular através dos quais penetram os vetores plasmídicos ligados à membrana através dos íons cálcio.
Em seguida, as células são plaqueadas em placas de Petri com meio seletivo e as colônias crescem sobre elas.
3.2.2) Eletroporação
A base do método de
eletroporação é o fato de que o campo elétrico pode danificar a membrana celular. Isto é, literalmente, a eletroporação é a criação de poros por um campo elétrico.
Primeiro, as bactérias e um vetor plasmídico são colocados em um tubo especial (cubeta). Em seguida, a cubeta é colocada em um dispositivo especial, que por uma fração de segundo cria uma diferença potencial de 0,1 a 10 kV, o que, por sua vez, leva a danos reversíveis nas membranas e os vetores podem penetrar livremente nas células. Após algum tempo, a cultura bacteriana é semeada em placas de Petri com meio seletivo.
Um requisito importante para o meio líquido no qual as células e o vetor estão localizados no momento da eletroporação é uma baixa concentração de íons: caso contrário, pode ocorrer um curto-circuito.
Posfácio
Cada colônia em uma placa é a descendente em proliferação de
uma única bactéria (ou seja, se vemos 100 colônias em uma placa, significa que havia inicialmente 100 bactérias viáveis, que começaram a se dividir).
Depois que as colônias aparecem nos pratos, o pesquisador seleciona uma das colônias e transfere as células da colônia para um novo meio nutritivo líquido seletivo. Depois que uma nova cultura de células "cresce", é novamente plantada em um novo copo com o mesmo meio nutritivo seletivo. Como resultado, obtemos um copo no qual apenas os descendentes de bactérias de uma única colônia crescem (ou seja, todos são clones de uma única bactéria). E somente essas células serão usadas no futuro para obter a proteína necessária em grandes volumes de meio nutritivo seletivo líquido.Bem, vamos resumir os resultados intermediários:- Nós sintetizamos o gene que precisamos;
- Insira-o em um vetor plasmídico adequado;
- Uma estirpe produtora adequada foi selecionada e transformada;
- Tem uma cultura de clones;
- Finalmente, transferimos parte desses clones para um grande volume de meio nutriente, a fim de obter a proteína necessária em um grande volume.
O próximo passo: isolar a proteína da biomassa resultante da forma mais pura possível. Mas esta é uma história completamente diferente.