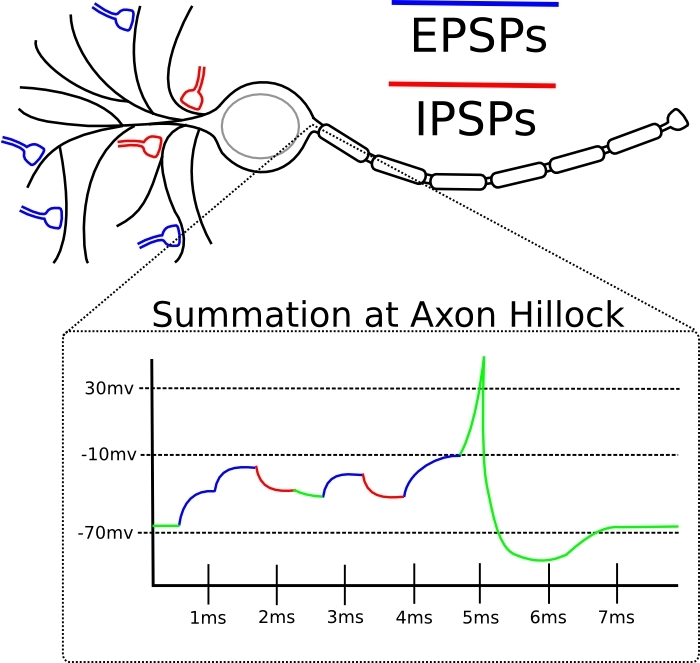
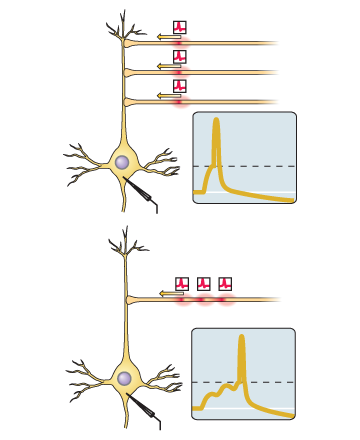
如果此突触的输入尖峰趋向于恰好在神经元本身生成尖峰之前,则突触被放大。如果此突触的输入尖峰趋于在神经元自身产生尖峰之后立即出现,则突触会减弱。
Source: https://habr.com/ru/post/zh-CN390385/More articles:首次接触小行星15年在线约会网站和应用程序的用户数量正在迅速增长Ubuntu平板电脑:“您所需的PC都在平板电脑中”你工作太多吗 休假无济于事2月14日-劳伦斯日:送给亲人的礼物权利人发明了将海盗转变为付费用户的新方法FRAM技术通讯质量-交通部的Android应用程序如何唤起人们对历史或战争游戏的兴趣智能银行木马可让您在自动柜员机上提取几乎无限量的资金All Articles