在我以前的出版物中 ,考虑了两个问题:分子生物学的基础知识以及基于质粒载体构建基因所需的构建体的方法。 现在,您需要弄清楚如何将质粒载体引入细菌细胞,即进行“转化”。 同时,我们将学习有关细菌膜的结构及其克服方法的知识,以及为什么某些细菌被称为主管细菌,所有这些与细菌毒素以及细菌对抗生素的抗性如何相关以及许多其他有趣的事实。
因此,我们手上有一个质粒载体,并且有一个适合该载体的细菌菌株,仍然需要将该载体引入细菌中。
在
上一篇文章中 ,提到
了弗雷德里克·格里菲斯(Frederick Griffith )的
工作 ,这清楚地证明了DNA可以自发渗透细菌细胞的事实。 细菌细胞吸收环境DNA分子的能力称为“能力”(这引起了很多笑话,即即使在体外单细胞也能胜任,而您的实验室同事却没有能力)。 如果吸收不是在某些人为创造的条件的影响下发生的,那么我们所说的是“自然能力”。 细菌内部如何发生类似的DNA转运?
1)细菌细胞膜的结构
首先,我们简要地考虑革兰氏阳性和革兰氏阴性细菌的膜结构。
革兰氏阴性细菌的外壳由两个脂质双层组成:外膜和内膜(内膜也称为血浆)。 在它们之间是周质空间,其中充满了执行重要功能的蛋白质和肽聚糖。
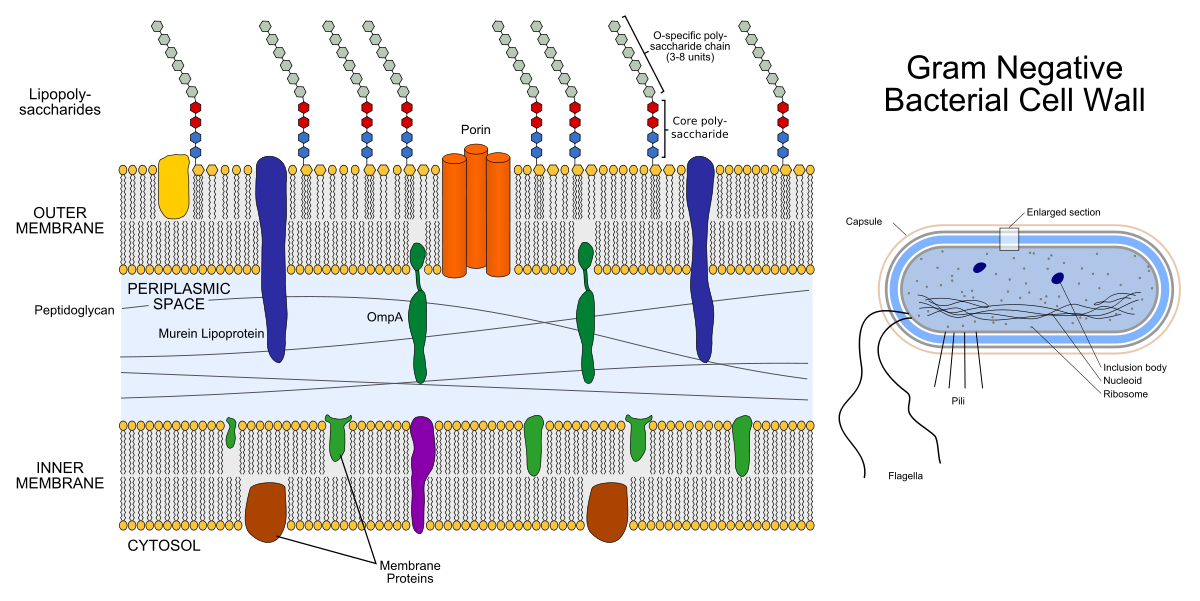 左边是革兰氏阴性细菌膜的结构。 膜内部的彩色小物件是膜蛋白,伸出的链是脂多糖。 右边是革兰氏阴性细菌本身的结构。
左边是革兰氏阴性细菌膜的结构。 膜内部的彩色小物件是膜蛋白,伸出的链是脂多糖。 右边是革兰氏阴性细菌本身的结构。肽聚糖是一种杂聚物,由共价交联的N-乙酰氨基葡萄糖(NAG)和N-乙酰氨基甲酸(NAM)组成。 原来是链....-NAG-NAM-NAG-NAM-NAG- .... 术语“杂聚物”中的“杂”是指该聚合物不是由相同的元素(如聚乙烯)组成,而是由两种不同的元素组成。 肽聚糖合成酶转肽酶是β-内酰胺类抗生素(青霉素类抗生素和头孢菌素类)的靶标:由于细菌分裂的过程,它们抑制了其活性,两个子细胞都没有足够的肽聚糖。 肽聚糖层也是溶菌酶的目标:它破坏NAG和NAM之间的共价键。 在医学上,溶菌酶被用作防腐剂,例如,它是某些用于治疗喉咙痛的药物的活性成分,在食品工业中,它可以用作防腐剂(食品添加剂E1105-这是溶菌酶)。
肽聚糖。 该图显示了与一个NAG(在左侧)和一个NAM(在右侧)相对应的部分,它们通过一个氧原子相互连接。 (对于那些在化学上不喜欢这种自由的人:这种“通过氧原子键”称为β-(1,4)-糖苷)。革兰氏阳性细菌的包膜仅由内膜(质膜),非常薄的周质空间和比革兰氏阴性细菌更厚的外部肽聚糖层组成。
除水以及小的非极性和疏水性分子(分子氧和氮,二氧化碳等)外,阻碍细菌细胞质中任何化合物的此类障碍是无法克服的。 这些化合物通过壳的渗透称为“被动迁移”(“简单扩散”),它总是沿着浓度梯度发生。 非极性分子能够自由通过膜,因为膜的内部容积也是非极性的(在膜内部,它们由脂肪酸的疏水残基组成),非极性化合物在非极性溶剂(酒精,丙酮,脂肪酸)中的溶解性很好。
显示了两层膜内部脂肪酸残基的疏水性(疏水性)的漫画图片。反过来,大极性分子(氨基酸,单糖,核苷酸),带电粒子(离子)和大分子(DNA,蛋白质)不能像那样穿透细胞膜,因为它们仅能很好地溶于极性溶剂(例如,水中) 。 那么DNA分子如何进入细菌细胞呢?
2)诱导化合物从环境进入细胞的机制
首先,我们有一个显而易见的事实,即细菌不能仅依靠水,氧气和二氧化碳来生存,因为它的生命还需要许多其他化合物。 因此,有一些方法可以将这些化合物输送到细胞中。 这种方法是“促进扩散”和“主动运输”。
2.1)光扩散
促进扩散的例子(尽管这种特殊情况相反,对细菌有害)是钾离子与缬氨霉素(一种由某些真菌合成的抗生素)结合通过细菌细胞膜的转运。 游离的缬霉素不能渗透到细胞中,因为极性基团暴露在外部。 钾离子也是“封闭的道路”。
 雕塑描绘了缬霉素和钾离子的复合物的三维结构(离子被描绘成中心的球形)。 它安装在俄罗斯科学院(莫斯科)M.Shemyakin和Yu.A.Ovchinnikov生物有机化学研究所入口的前面,其中建立了缬霉素的抗菌活性机理及其与钾离子的复合物的三维结构。
雕塑描绘了缬霉素和钾离子的复合物的三维结构(离子被描绘成中心的球形)。 它安装在俄罗斯科学院(莫斯科)M.Shemyakin和Yu.A.Ovchinnikov生物有机化学研究所入口的前面,其中建立了缬霉素的抗菌活性机理及其与钾离子的复合物的三维结构。但是,缬氨霉素与钾离子形成复合物后,它改变了三维结构,使其整个外表面变为疏水性(非极性),而所有亲水基团都隐藏在内部。 在这种情况下,钾离子隐藏在缬霉素分子内部;因此,缬霉素+钾离子复合物可以穿透膜。 该复合物可以在细胞内外随时都分解为游离的缬霉素和离子,但是通常促进离子的扩散导致它们在细胞内和细胞外空间的浓度均衡,并且细胞控制离子的运输是有原因的:它们的高浓度致命的 瓦里霉素的抗菌作用正是基于此。
与钾离子复合的真菌抗生素缬氨霉素的示意图。另一个例子是不受控制的孔,它们是大的跨膜蛋白,可以使某些物质根据梯度自由通过膜。 这种跨膜孔的一个例子是
金黄色葡萄球菌 α-毒素(毒素的另一个名称是α-溶血素。它由“血红素”和“赖氨酸”部分组成。“血红素”表示红细胞;“赖氨酸”表示来自“裂解”一词,即“细胞死亡”)
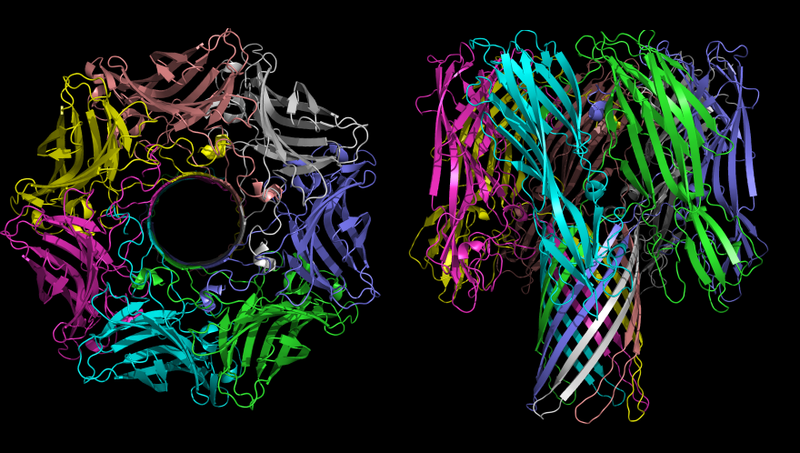 α-溶血素低聚物的三维结构-金黄色葡萄球菌的毒素。 在左侧的图片中,时间清晰可见。
α-溶血素低聚物的三维结构-金黄色葡萄球菌的毒素。 在左侧的图片中,时间清晰可见。α-溶血素在致病过程中的主要功能是在细胞膜中形成不受控制的通道,根据环境条件,通道可以通过单价离子,钙离子或ATP。 如上所述,离子失衡具有极强的毒性,并且ATP
从细胞中的运输是有害的,因为ATP是任何细胞几乎所有化学过程中必不可少的环节。
2.2)主动运输
从上面的例子可以看出,促进扩散不需要能量。 相反,主动转运是细胞吸收或释放某种东西,而每次转移都需要消耗能量。 这可以是大极性分子或离子的传输,离子的传输既可以沿浓度梯度进行,也可以沿浓度梯度进行。 当然,主动转运是受控过程,其中涉及整套细胞内和跨膜蛋白。 因此,细胞首先需要花费资源来合成所有这些蛋白质,然后再次需要花费能量来使用该转运系统。
主动转运的一个例子是将能量和疏水性和两亲性化合物从细胞转运回环境(外排)的能量依赖系统。
抗生素从细菌细胞向外部主动转运的系统,该系统由形成单个跨膜复合物的三种蛋白质组成。 该图反映了一个事实,即外排转运蛋白(紫罗兰)可以直接在细胞质和膜中“捕获”药物分子。该系统特别令人感兴趣,因为它会导致某些细菌对抗生素产生多重耐药性:在严重破坏细菌之前,细胞已将其清除。 此外,
铜绿假单胞菌铜绿假单胞菌的多药耐药病例中有一半以上与抗生素的主动转运有关。 更重要的是,已经发现了“新型”抗生素的主动外排系统,可用于抵抗对常用抗菌药物产生耐药性的细菌。 例如,在经典抗生素无效的情况下使用的药物
利奈唑胺被基于AcrAB和TolC蛋白的外排系统抛弃了。 另一个有趣的事实是,在不需要细菌的情况下,智能细菌不会在外排系统蛋白质的大规模合成中浪费能量。 但是,一旦出现对细胞有害的物质,合成就会被迅速激活,细菌开始主动抵抗。 顺便说一句,外排不是保护细菌免受抗生素侵害的唯一机制,我还将写一篇有关其他方法的文章。
利奈唑胺的结构是一种抗革兰氏阳性菌耐药菌株的抗生素。3)细菌细胞的能力
因此,我们研究了化合物的跨膜转运机制,这些机制如果没有外部援助就无法穿透该屏障。 在自然条件下,由于细胞内和跨膜蛋白的整个光谱的共同作用,DNA仅通过主动运输才能进入细菌。 细胞从外界吸收DNA的未诱导能力称为“自然能力”。
3.1)细菌细胞的自然能力
细菌自然能力的出现的最确凿的假设是这种机制允许细菌在极端条件下存活(细菌的极端条件称为“应激”),这一假设是细菌的自然能力。 事实是,压力通常会导致细菌的“染色体” DNA受损,如果细胞想要存活,则需要以某种方式“修复”其“染色体”(这种“修复”称为“修复”)。 此外,众所周知,如果将与细胞自身DNA某些部分相似的DNA序列引入细胞,那么很可能会将这一“新”片段整合到其基因组中。 在压力条件下会发生什么? 一些细胞死亡,成为仍然为生命而战的兄弟的DNA捐赠者。 确实,在同一细菌培养物中,代表基因组很可能非常相似,因此,存活的细菌可以使用死者亲属的DNA修复其基因组。
此外,在舒适的细胞培养条件下,只有一小部分具有自然能力,而感受态细胞在其总数中所占的比例最大,那么培养就处于一定的生长阶段-对
数期。
 一段时间内培养物中典型的细菌图。 对数期是生长期,然后是平稳期和灭绝期,在此阶段,细胞已经缺乏营养,毒素蓄积。
一段时间内培养物中典型的细菌图。 对数期是生长期,然后是平稳期和灭绝期,在此阶段,细胞已经缺乏营养,毒素蓄积。当然,研究人员希望使质粒载体转移到细胞中的过程尽可能有效:转化过程越有效,为了保证良好的结果,所需的质粒载体就越少。 因此,在实验室条件下,它们通常不依赖细胞本身,而是创建具有人工能力的细胞。
3.2)细菌细胞的人工能力
建立细菌人工能力的主要方法有两种:用二价离子处理细胞培养物,然后进行短期加热和所谓的电穿孔。
3.2.1)用二价离子处理细胞培养物,然后短期加热。
通常,在实验室中,有一台温度约为-80摄氏度的冰箱(有时为纪念我们国家的一家制造公司而被称为“开尔文发生器”),该冰箱被处于对
数期的装有试管的试管堵塞了。 这样做是为了避免浪费时间在每次需要它们时都创建它们(并且要花费很多时间,因此它不会比一天中更快地工作)。 为了使细胞在储存过程中不会因冷冻而死亡,可将冷冻保护剂添加到培养基中:甘油,丙二醇或二甲基亚砜。
图片作者认为实验室冰箱。当研究人员需要转化特定菌株时,他从开孔器中取出细胞,并将试管放入冰中。 然后,在生长的对
数期将二价阳离子和我们需要的质粒载体添加到细胞中。 试管不得离开冰块。 这种阳离子的典型供体是氯化钙(CaCl2),在这种情况下,我们的二价阳离子是Ca2 +。 关于Ca2 +如何帮助DNA渗透细菌没有明确的解释:
- 一些消息来源声称,高浓度的Ca2 +会引起细胞膜间隙和DNA损伤,而我们早些时候发现DNA损伤是细胞胜任能力的信号。
- 根据其他来源,这些离子充当将质粒载体附着至细胞的“桥”。 事实是,带正电的离子会同时附着在细菌外膜上的多糖的带负电的基团和质粒DNA的带负电的基团上。 如果培养基中没有离子,那么DNA将被细胞排斥,这对研究人员完全没有用。
两种假设都是正确的。
推测Ca2 +离子参与转化的机理。 首先,离子附着到带负电荷的DNA组(黄色圆圈)和锚定在膜上的多糖(黄色圆圈)上。 然后,由于离子的作用,形成了一种桥:“多糖(-)”-“钙离子(2 +)”-“ DNA(-)”,它将DNA固定在细胞表面。在接近零温度的细菌温育后,产生了所谓的热激-将培养物短期加热至42度。 通常,热处理的持续时间为30-45秒,此后带有细菌的试管返回冰浴(如果加热时间更长,细胞可能会死亡)。 造成热休克有效的原因可能是相同的机制:高温会损坏DNA和蛋白质(这会引起竞争),还会在细胞膜上形成“孔”,通过钙离子附着在膜上的质粒载体会穿透这些孔。
然后将细胞铺在具有选择性培养基的培养皿上,并在其上生长菌落。
3.2.2)电穿孔
电穿孔方法的基础是电场会损坏细胞膜的事实。 即,从字面上看,电穿孔是由电场产生的孔。
首先,将细菌和质粒载体置于特殊的试管(比色皿)中。 然后将比色皿放置在特殊的设备中,该设备会在瞬间产生0.1-10 kV的电势差,进而导致对膜的可逆破坏,载体可以自由地渗透到细胞中。 一段时间后,用选择性培养基将细菌培养物接种到培养皿中。
电穿孔时细胞和载体位于其中的液体介质的一个重要要求是离子浓度低:否则,可能会发生短路。
后记
平板上的每个菌落都是
一种细菌的繁殖后代(也就是说,如果我们在平板上看到100个菌落,则意味着最初有100个活菌,然后开始分裂)。
菌落出现在盘子上后,研究人员选择其中一个菌落,并将细胞从菌落转移到新的选择性液体营养培养基中。新的细胞培养物“长大”后,将其再次种植在具有相同选择性营养培养基的新杯中。结果,我们得到了一个杯子,只有单个菌落的细菌后代会在上面生长(也就是说,实际上它们都是单个细菌的克隆)。将来,只有这些细胞才能用于在大量液体选择性营养培养基中获得我们所需的蛋白质。好吧,让我们总结一下中间结果:- 我们合成了所需的基因;
- 将其插入合适的质粒载体中;
- 选择合适的生产菌株并转化。
- 有克隆文化;
- , .
: , . .