神经发生是成熟的中枢神经系统(CNS)中新神经细胞形成的多阶段过程,这是其适应功能。
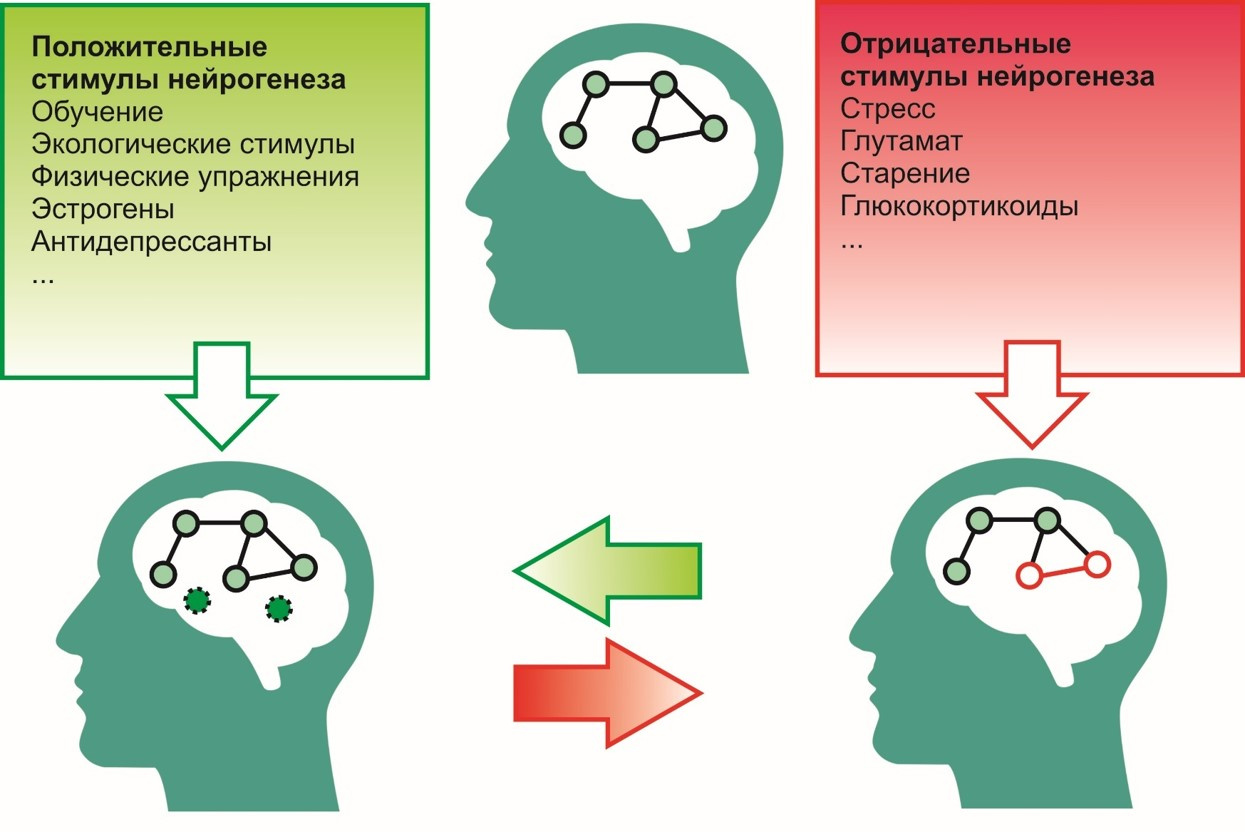
积极奖励措施 促进神经发生过程的诱因包括:
- 学习过程
- 生态环境(有利环境),
- 体育锻炼(例如跑步)
- 抗抑郁药
- 雌激素等
否定-
- 压力
- 谷氨酸在中枢神经系统中的过度活动,
- 暴露于糖皮质激素(皮质醇-压力激素),
- 衰老等。
多年来在中枢神经生物学领域,有一些中枢教条不允许神经发生。
 成熟脊椎动物的大脑中不存在神经发生的概念基于以下四个原则:
成熟脊椎动物的大脑中不存在神经发生的概念基于以下四个原则:- 临床。 患有以中枢神经系统病变为主的神经病理学疾病的患者不会经历功能恢复。 脑血管疾病,外伤性病变和神经退行性疾病,例如帕金森氏症和阿尔茨海默氏病,是中枢神经系统的病理,患者的病情不断恶化。 通常,各种治疗方法只能消除症状,而不能阻止疾病的发展。
- 功能性。 它基于以下事实:中枢神经系统控制着许多复杂的过程:情绪,运动,反射等的调节。 管理如此复杂的过程需要极其精确和微调。 人们相信,新神经细胞的形成,它们的分化和迁移可能破坏现有神经通路的结构和功能组织,并破坏中枢神经系统。
- 与记忆和学习理论相关。 长期以来,人们认为“记忆”是从训练过程中形成的神经网络中提取信息。 在这种情况下,新神经细胞的形成被认为是与记忆不相容的事件。
- 技术和实验。 在成熟的中枢神经系统中缺乏发现神经发生和神经干细胞的技术能力。
关于成熟大脑神经发生的思想形成阶段成年(或成熟)大脑的主要神经源性区域是海马的齿状回和侧脑室的脑室下区域。 区域显示在图中。
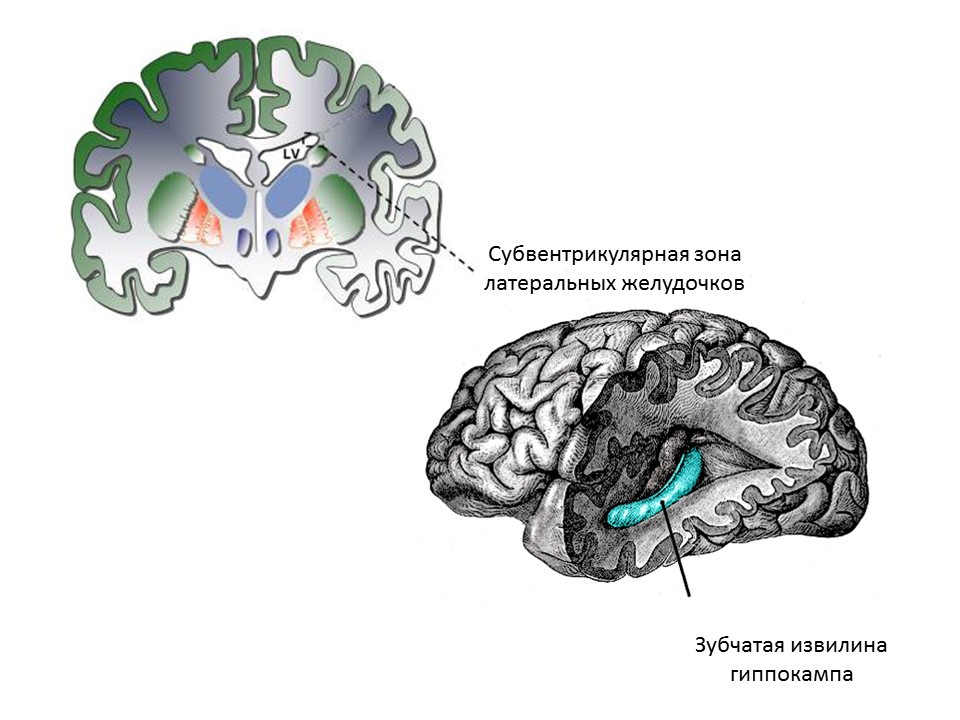 成熟大脑海马齿状回中神经发生的阶段
成熟大脑海马齿状回中神经发生的阶段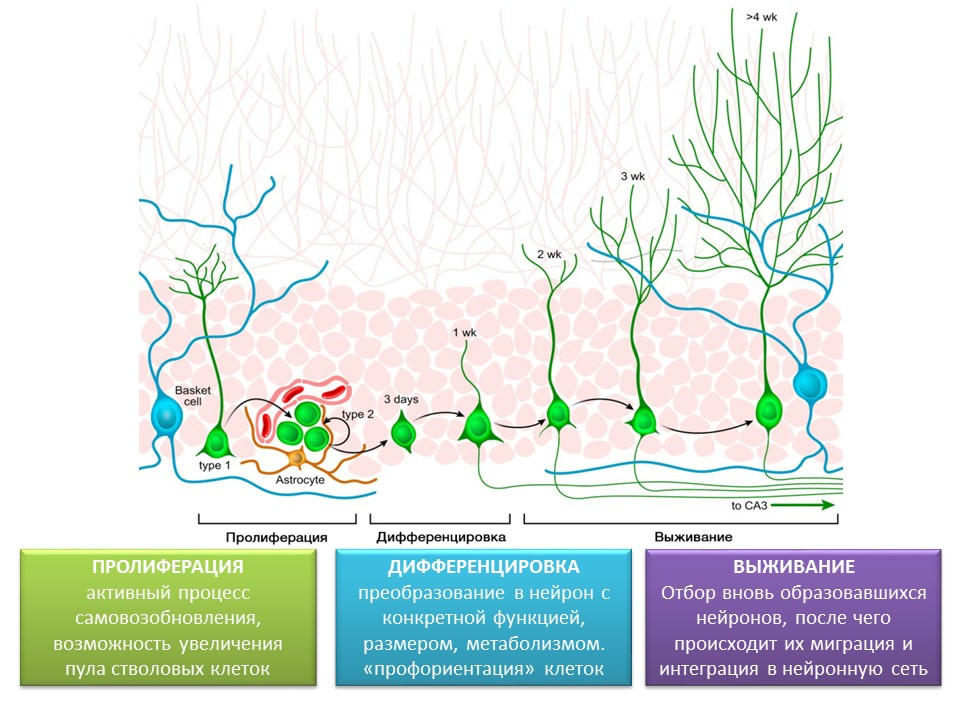
增殖是自我更新的活跃过程,是增加干细胞库的可能性。
分化-转换为具有特定功能,大小和代谢的神经元。 实际上,是未来神经细胞的“职业指导”。
生存是选择新形成的神经元,然后它们迁移并整合到神经网络中。 神经细胞的选择是一部分形成的神经元的凋亡(程序性死亡)。 其余的细胞迁移到大脑的相应区域并整合到神经网络中。
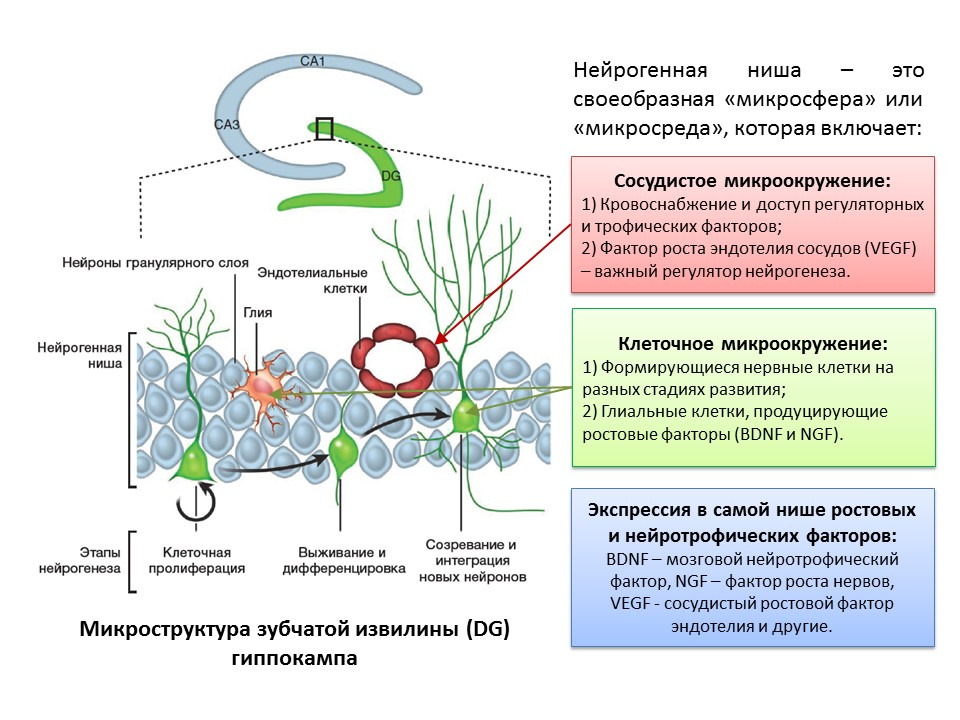
术语“神经发生”与“神经源利基”的概念密不可分。 “神经源性利基”本身就是其中发生神经发生过程的“微球”。 神经源性利基包括:
- 血管微环境,其首先为利基自身提供血液并确保各种调节分子的传递(在靠近血管的位置形成并转化干细胞,其次,血管内皮生长因子(VEGF)不仅是重要的调节因子在血管生成(已存在的血管系统中新血管的生长)和血管生成(胚胎血管系统的形成)过程中,还直接调控神经发生;
- 细胞微环境,包括处于发育各个阶段的神经细胞和执行营养功能的神经胶质细胞(它们产生生长因子,例如脑神经营养因子BDNF和神经生长因子NGF);
- 表达在利基增长和调控因素。 除了神经胶质细胞和利基的其他成熟结构外,干细胞还能够产生调节因子和生长因子,从而进行自我调节(自分泌调节)。
使用特定标记物评估神经发生过程
一个值得关注的重要而有趣的问题是如何检测成熟哺乳动物的大脑组织中的神经发生问题。
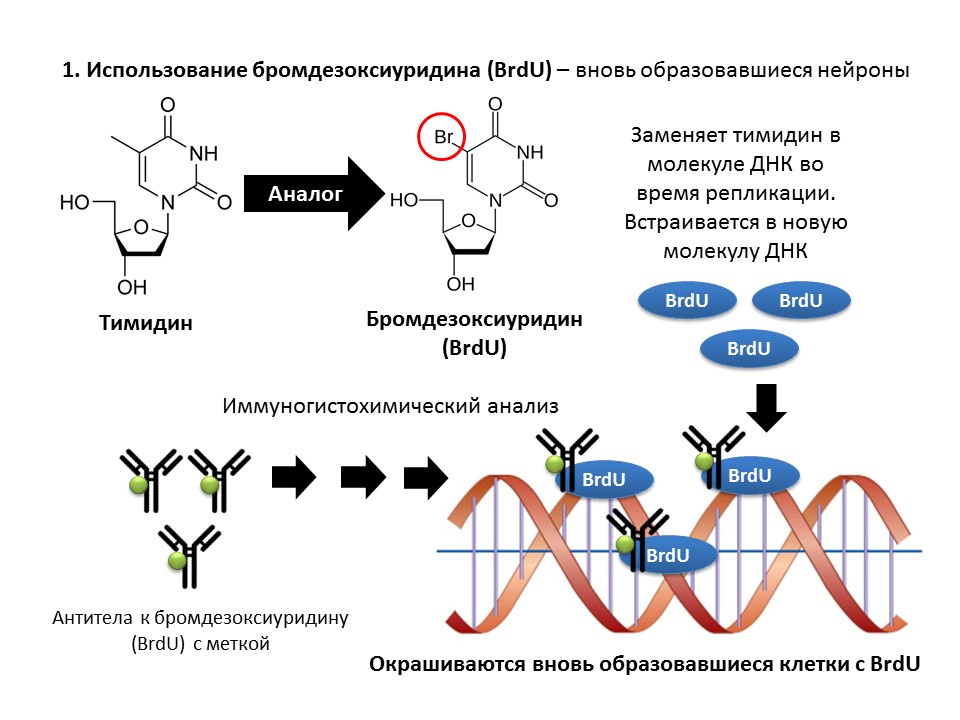
溴脱氧尿苷(BrdU)是胸苷的结构类似物-胸腺嘧啶脱氧核苷是DNA分子的组成部分。 将BrdU引入人体后,它会插入到分裂细胞的DNA中而不是胸腺嘧啶核苷中,从而提供了检测新形成的细胞并将其与“旧”细胞分离的机会。 获得脑组织样本后,将它们用针对BrdU的抗体(含有荧光标记的抗体)处理,该抗体通过抗原抗体免疫化学反应机制与BrdU结合,并能够对BrdU进行比色测定。 因此,在微量制备中,可以定量标记有BrdU的细胞,即所谓的BrdU阳性细胞。
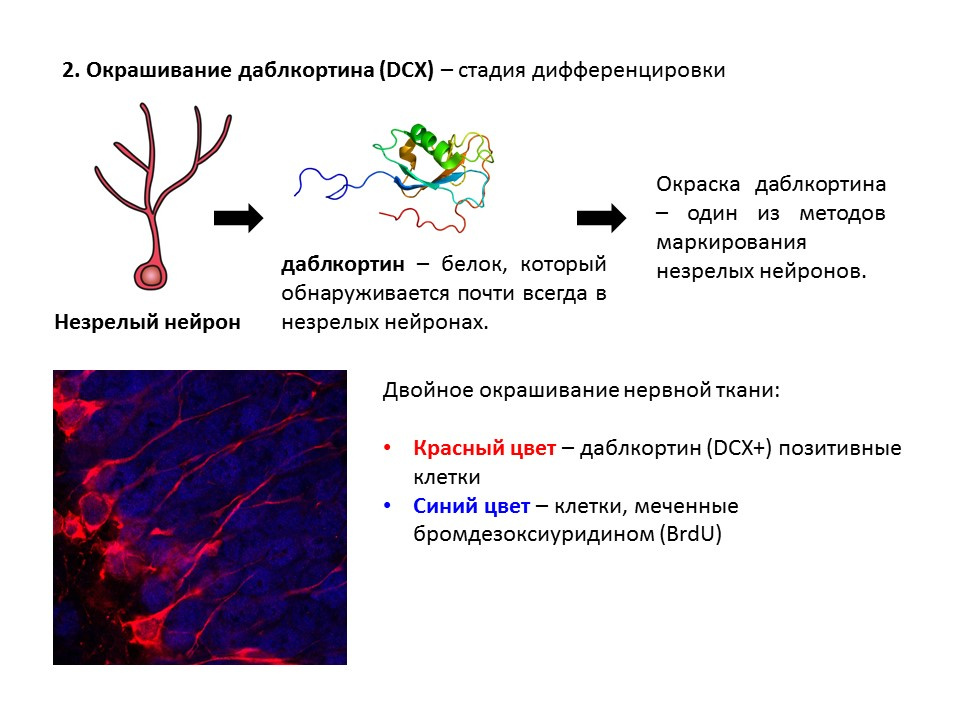
Doublecortin是一种蛋白质,几乎总是存在于未成熟的神经元中,并且可以被检测到。
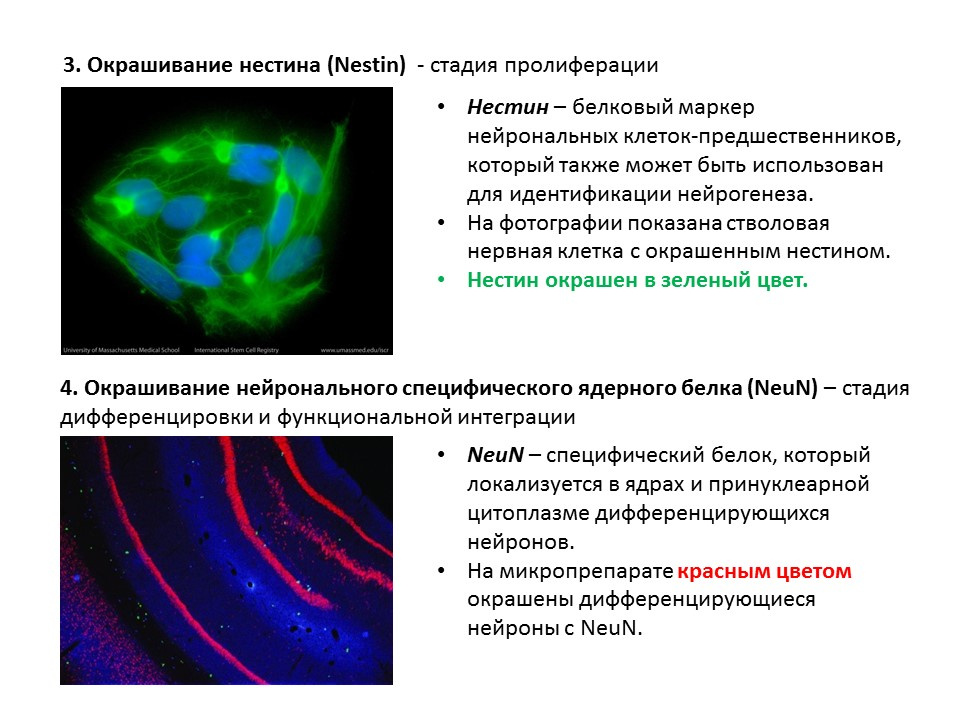
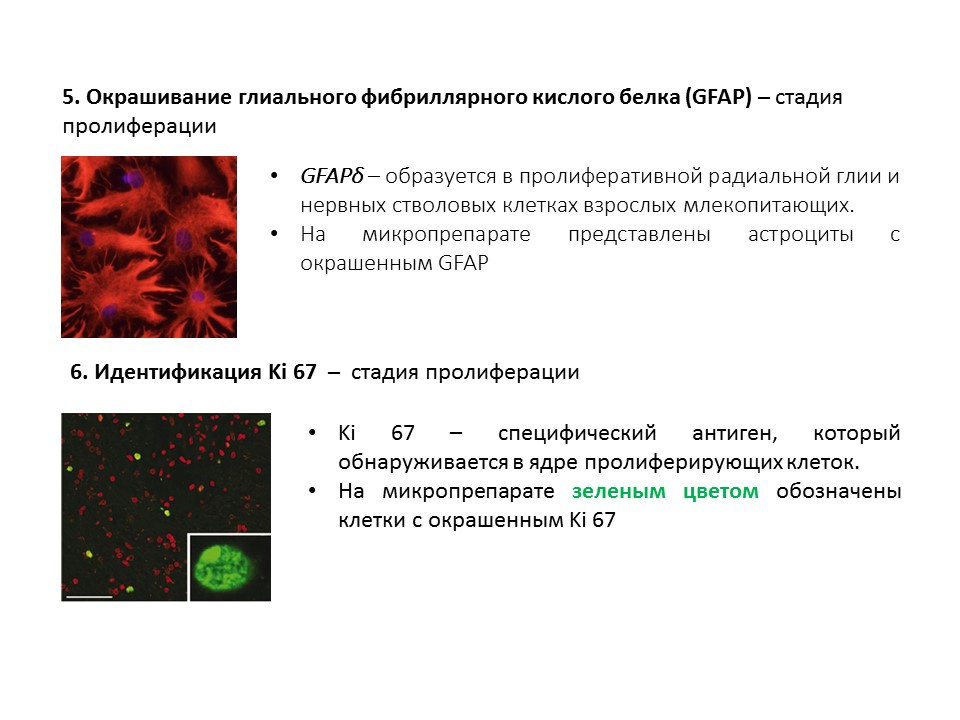 用于检测神经发生各个阶段的主要方法的摘要图,总结了较早提出的信息。
用于检测神经发生各个阶段的主要方法的摘要图,总结了较早提出的信息。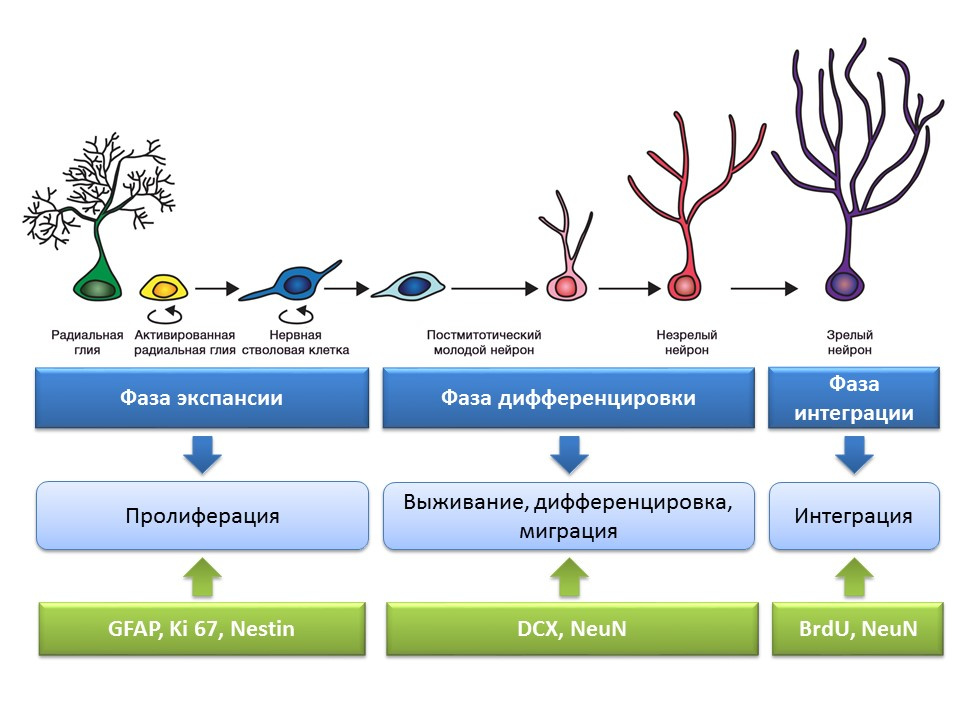
成熟大脑中神经发生的调节
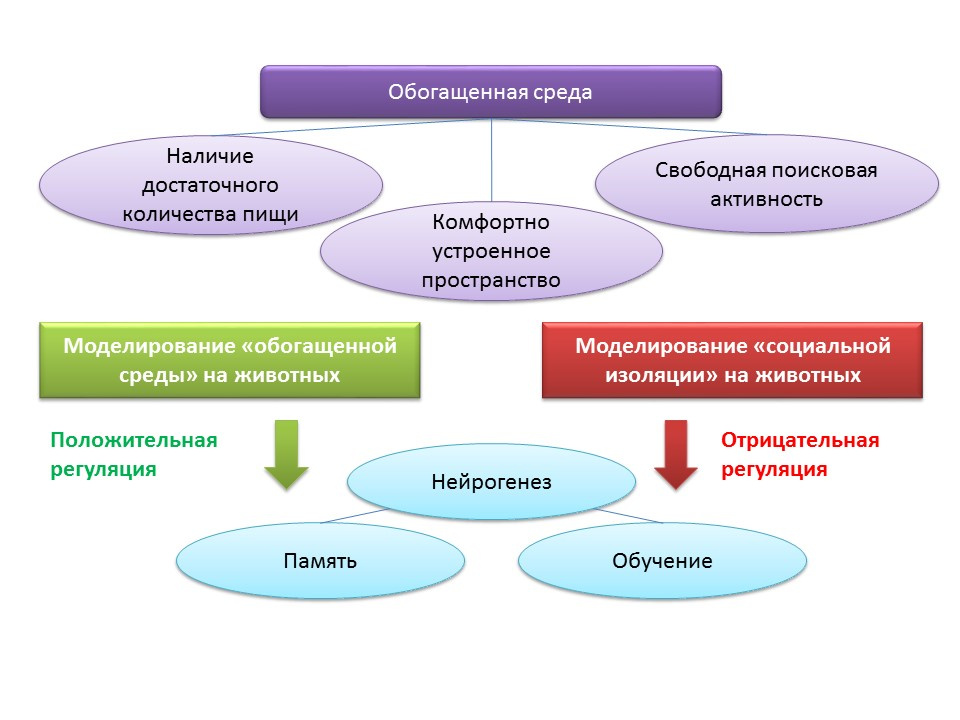
调节神经发生的环境因素与“丰富环境”的概念有关。 术语“丰富的环境”本身包括有利的生活环境,即存在足够的食物,舒适的空间和免费搜索活动的机会。
 DOI:10.1002 / hipo.22218DOI:10.1038 / 386493a0DOI:10.1016 / j.brainres.2011.08.00.007DOI:10.1016 / j.neuroscience.2011.10.040PMID:9547229
DOI:10.1002 / hipo.22218DOI:10.1038 / 386493a0DOI:10.1016 / j.brainres.2011.08.00.007DOI:10.1016 / j.neuroscience.2011.10.040PMID:9547229在动物实验中,发现留在“丰富的环境”中对神经发生具有积极作用:生长因子和神经营养蛋白的产生,增殖细胞的数量及其存活率增加。 神经发生的增加与动物认知功能的改善有关(主要与学习和记忆有关)。
将实验性C57BL / 6J小鼠在生物站(在林间空地)的条件下在一个受保护的空间中饲养2个月,对照组在标准实验室条件下进行饲养。 在实验组的动物中,与对照组相比,注意到兴奋性和抑制性突触的数目增加。 该实验表明环境条件可以有利地影响置于丰富环境中的啮齿动物的中枢神经系统的形态和功能。
相反,在实验室中将动物置于社会孤立的条件下,充当了神经发生的负调节器。
此外,在脑缺血后将动物置于“丰富的环境”中有助于增强大脑神经源性区域的再生过程。
这些研究的结果并不是什么特别的东西,因为在现代医学中使用康复课程和健康假期。
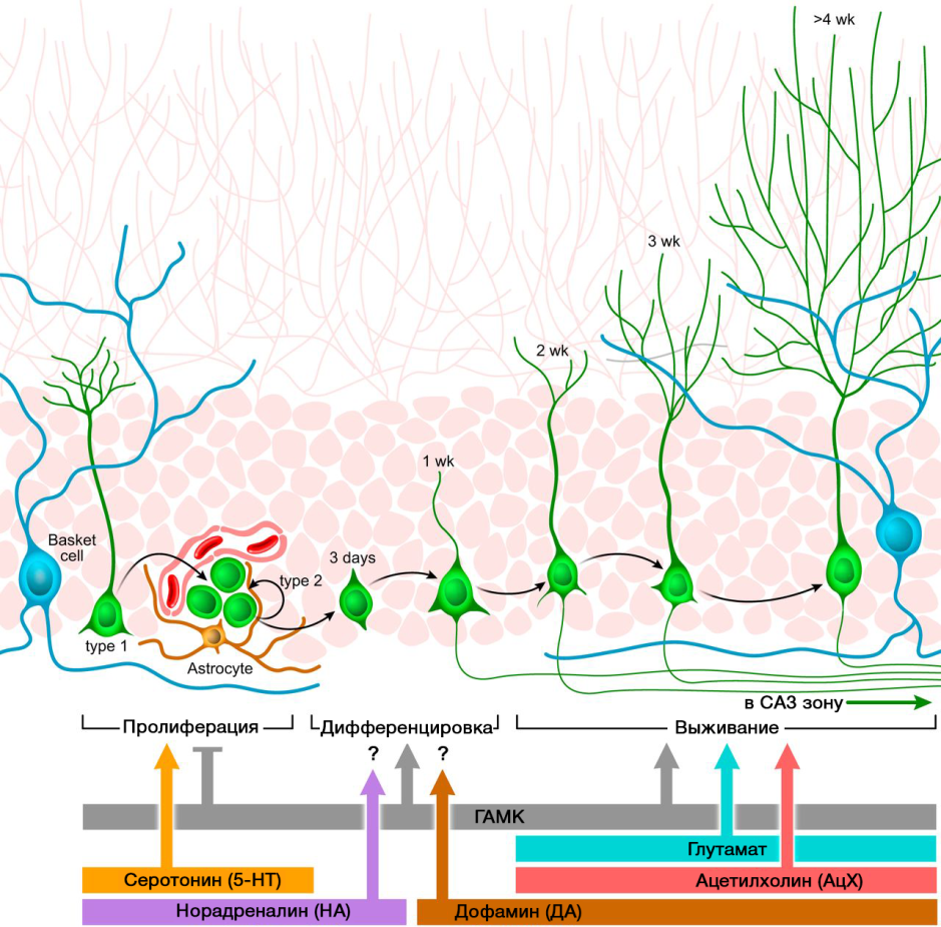
中枢神经系统神经递质在调节神经发生中起重要作用。 图片显示了这种调节的摘要图。
 DOI:10.1152 / physrev.00004.2014
DOI:10.1152 / physrev.00004.2014所提出的中枢神经系统神经递质在神经发生过程中的调节作用与中枢神经系统各种疾病中与神经发生水平变化有关的这些分子的缺乏或过量有关。
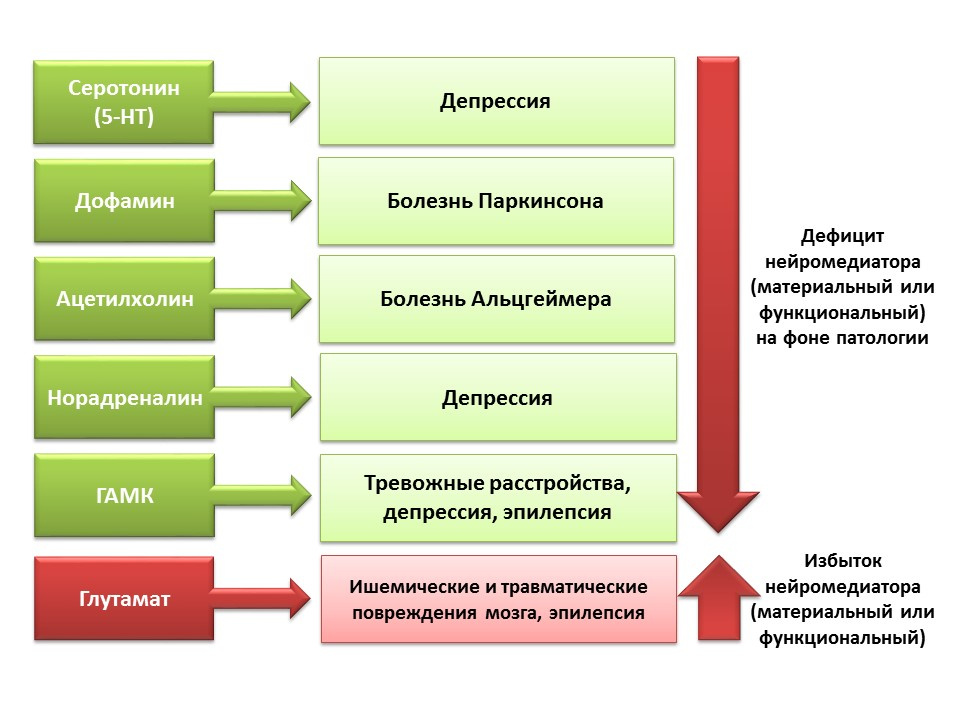
由于可能会对中枢神经系统的神经递质系统进行药理学操作,因此我们可以评估神经递质对成熟大脑神经发生各个阶段的贡献。
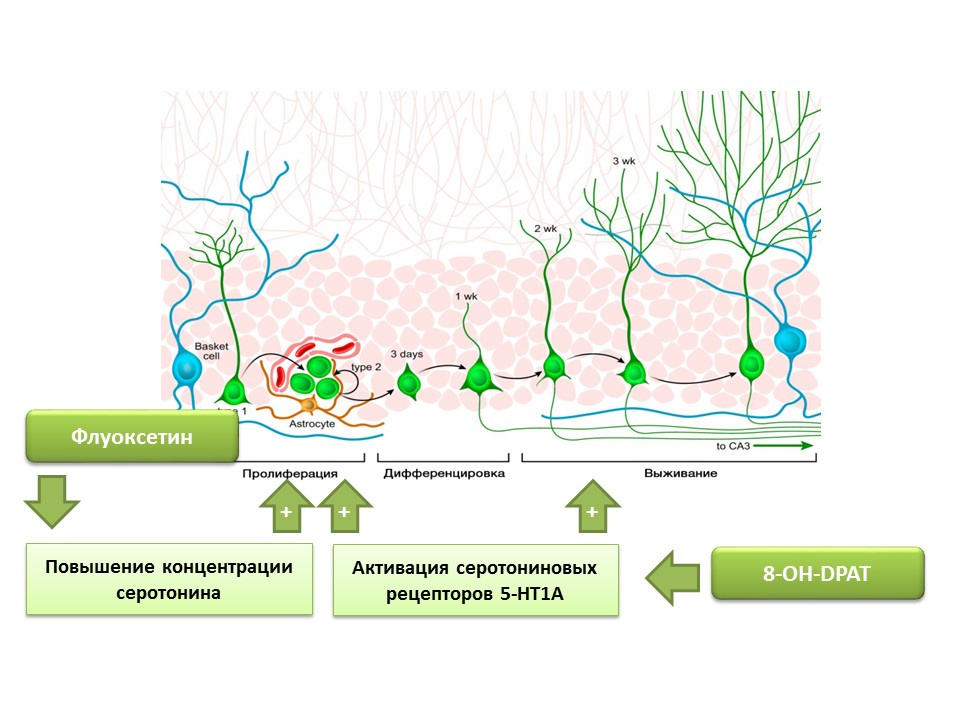
已确定氟西汀(一种选择性5-羟色胺再摄取抑制剂的抗抑郁药)可以增加中枢神经系统中5-羟色胺的浓度,增强啮齿类和灵长类动物海马齿状回中神经干细胞的增殖。 后来,在干细胞自身上检测到了血清素亚型1A受体(5-HT1A受体),这与8-OH-DPAT(这些受体的选择性激活剂)物质刺激实验动物(小鼠和小鼠)海马齿状回中新神经元的增殖和存活的能力是一致的。大鼠)。
灵长类动物暴露于慢性压力(社会隔离)中,此后评估了抑郁样和焦虑行为以及海马神经发生(死后)。 慢性应激会导致神经发生的减少,并伴有行为缺陷(增加抑郁症和焦虑行为)。 用氟西汀(一种抗抑郁药,通过抑制其再摄取来增加中枢神经系统中5-羟色胺的浓度)的治疗刺激了神经生成,并预防了抑郁症样和焦虑行为。
DOI:10.1371 / journal.pone.0017600
另一个实验证实了血清素在调节成人神经发生中的作用。 用8-OH-DPAT长期刺激大鼠5-HT1A血清素受体,可导致神经干细胞增殖增加,海马齿状回和脑室下区的分化神经元存活,并增加胶质发生。
DOI:10.1016 / j.euroneuro.2009.11.00.00
提出的实验证实了5-羟色胺在成人大脑神经发生过程中的重要调节作用。
 DOI:10.4161 / cc.8.18.9512DOI:10.1016 / j.neuropharm.2011.01.026
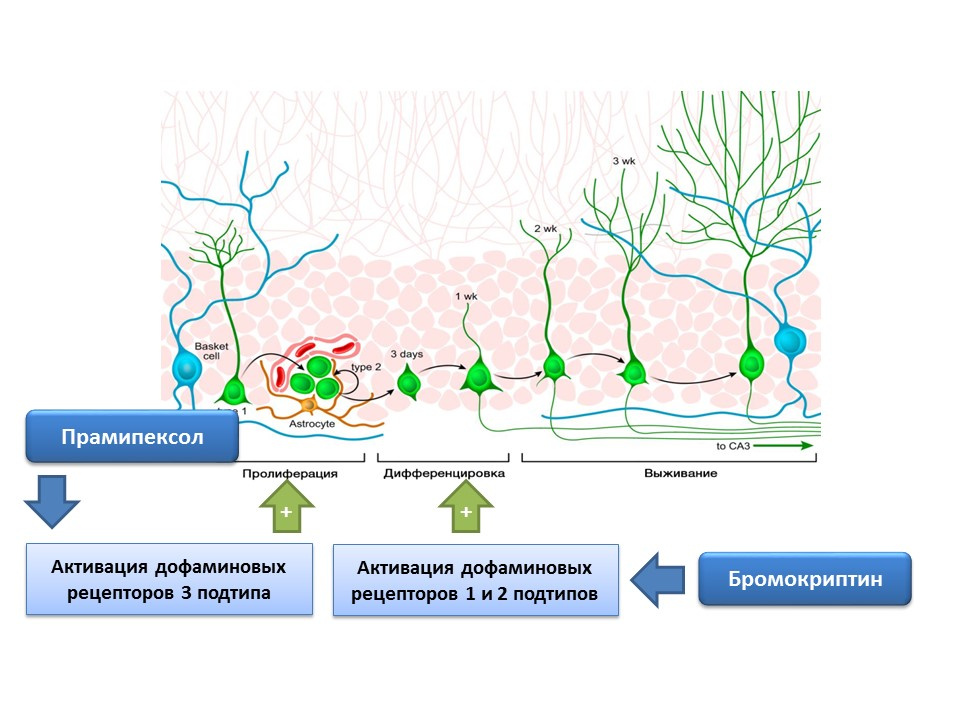
DOI:10.4161 / cc.8.18.9512DOI:10.1016 / j.neuropharm.2011.01.026多巴胺还积极参与神经发生的调节。 已经显示,用溴隐亭激活1型和2型多巴胺受体(D1和D2)会增强神经干细胞的分化过程,而用普拉克索刺激D3受体会导致增殖增加。
普拉克索是一种对D3受体的亲和力比D2大的多巴胺受体激动剂,用于帕金森氏病,可有效消除该疾病的症状。 另外,该药物具有神经保护活性。 在实验中,评估了普拉克索对小鼠细胞培养物中神经发生的影响。 结果显示,用普拉克索处理细胞培养物会导致神经球(新形成的干细胞簇)和含有双皮质素的细胞(未成熟神经元)的大小增加。 多巴胺受体阻滞剂-U99194A和舒必利消除了普拉克索对神经发生的刺激作用。 另外,已显示普拉克索增强了BDNF的释放。 提出的结果表明普拉克索在帕金森氏病中的有效性不仅与功能性多巴胺缺乏的补充有关,而且与对神经发生的刺激作用有关,并且还表明多巴胺在神经干细胞增殖阶段具有重要的调节作用。
DOI:10.1016 / j.neuropharm.2011.01.026
通过实验证实了神经干细胞上亚型1和2的多巴胺受体的存在,并且还显示了多巴胺在神经细胞培养物中增殖和分化中的重要调节作用。 特别地,在神经毒素MPTP损伤(在细胞培养中模拟帕金森氏病)的条件下,用溴隐亭(多巴胺受体D1和D2亚型的刺激物)处理脑室下区的细胞培养物,会导致细胞增殖和分化增加。
DOI:10.4161 / cc.8.18.9512
去甲肾上腺素能够通过激活β-3-肾上腺素能受体来增加神经干细胞前体的数量。 使用选择性β-3-肾上腺素受体激动剂(活化剂)-物质BRL37344显示了这种效果。
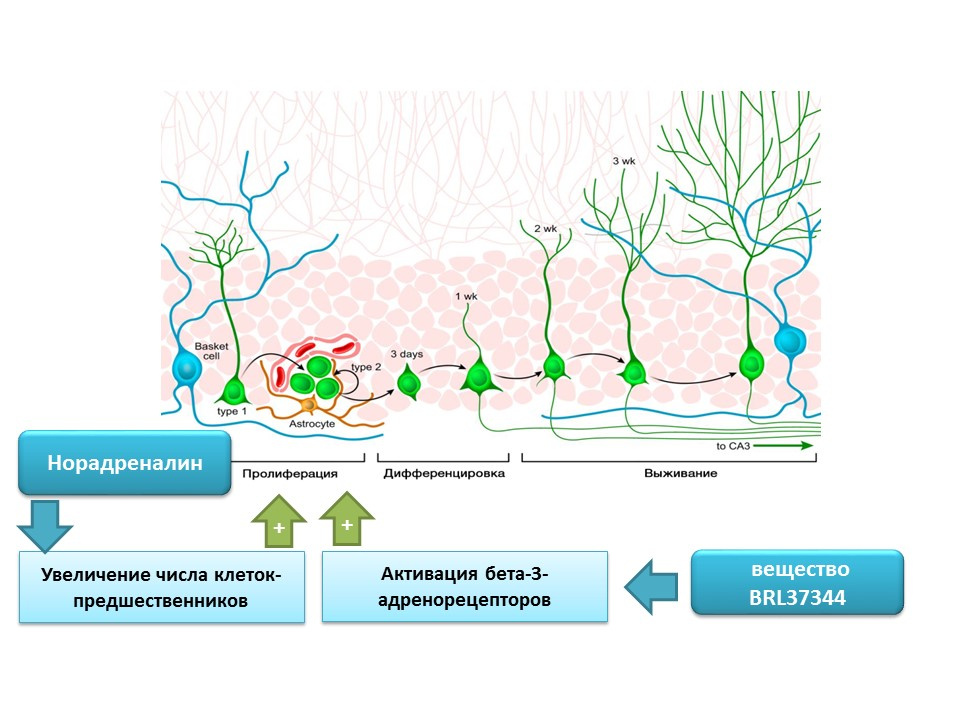
去甲肾上腺素在成熟神经发生过程中的调节作用已在一项全面的体外研究(在细胞培养中)在体内(在小鼠中)进行了评估。 向细胞培养物中添加去甲肾上腺素会导致神经球大小的增加(神经干细胞的积累)。 选择性β-3-肾上腺素受体激动剂的全身给药导致小鼠海马神经干细胞增殖增加。 这些研究证实了未成熟神经细胞上肾上腺素能受体的存在,并且还表明了去甲肾上腺素的调节作用,这显然是通过β-3肾上腺素能受体实现的。
DOI:10.1523 / JNEUROSCI.3780-09-09如前所述,神经干细胞本身能够产生调节因子。 这些因素之一是γ-氨基丁酸(GABA)-中枢神经系统的主要抑制介质。 特别是,在鼠神经细胞前体培养物中,神经干细胞增强了GABA的合成,而GABA的合成又通过激活放射状神经胶质(给细胞增加了干细胞)而增强了增殖,并在实验中促进了神经球大小的增加(干细胞蓄积)。
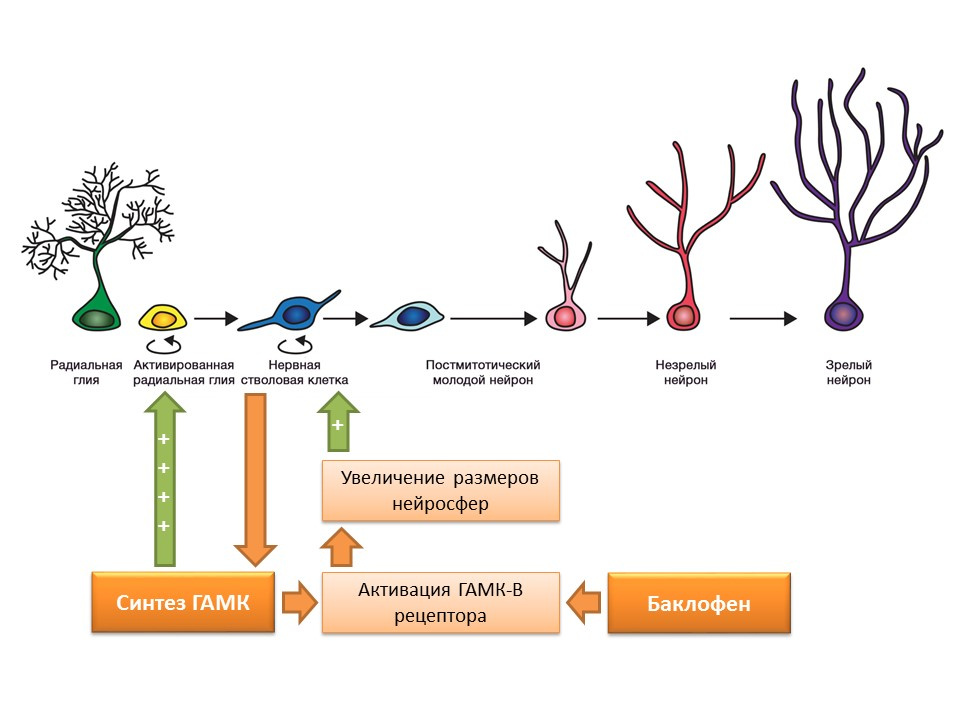
发现GABA的这种作用是通过GABA-B受体实现的。 该研究使用了选择性GABA-B受体激动剂-巴氯芬。
DOI:10.1002 / jcp.21422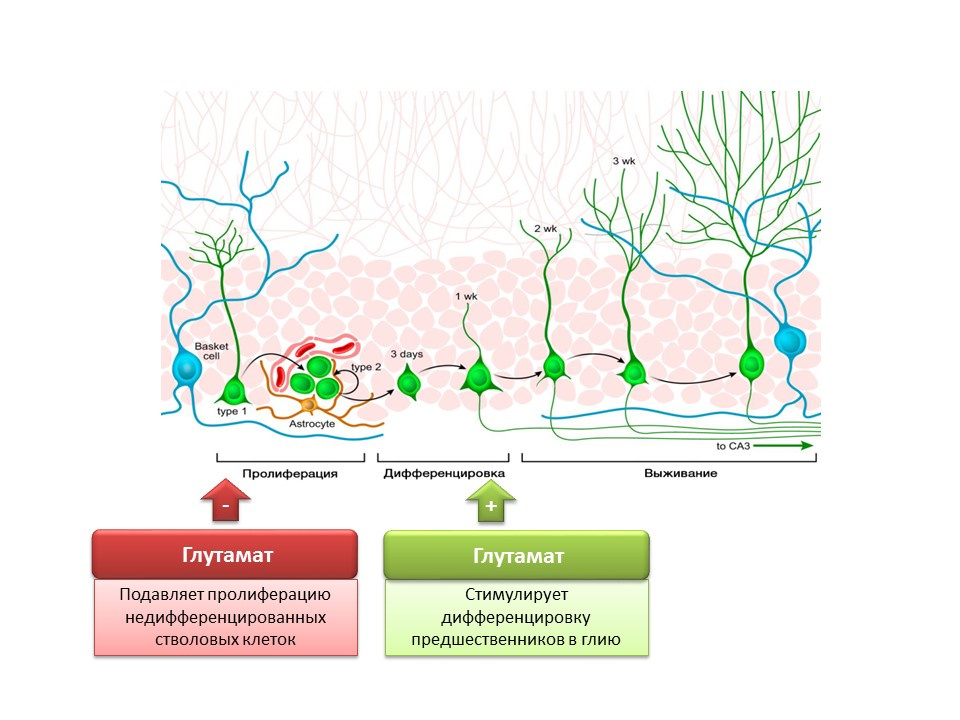 Nakamichi N,Takarada T,YonedaY。由γ-氨基丁酸和谷氨酸信号介导的神经发生。 J.Pharmacol Sci。 2009; 110(2):133-49
Nakamichi N,Takarada T,YonedaY。由γ-氨基丁酸和谷氨酸信号介导的神经发生。 J.Pharmacol Sci。 2009; 110(2):133-49主要兴奋介质在中枢神经系统中的作用是双重的。 一方面,谷氨酸抑制未分化干细胞的增殖。 另一方面,它刺激前体分化为神经胶质系。 认为一定比例的谷氨酸和GABA对于调节神经干细胞向某些类型的神经元的分化是必需的。 例如,如前所述,谷氨酸促进神经胶质细胞(星形胶质细胞)的形成,而GABA促进合成GABA本身的神经元(GABA能神经元)的形成。
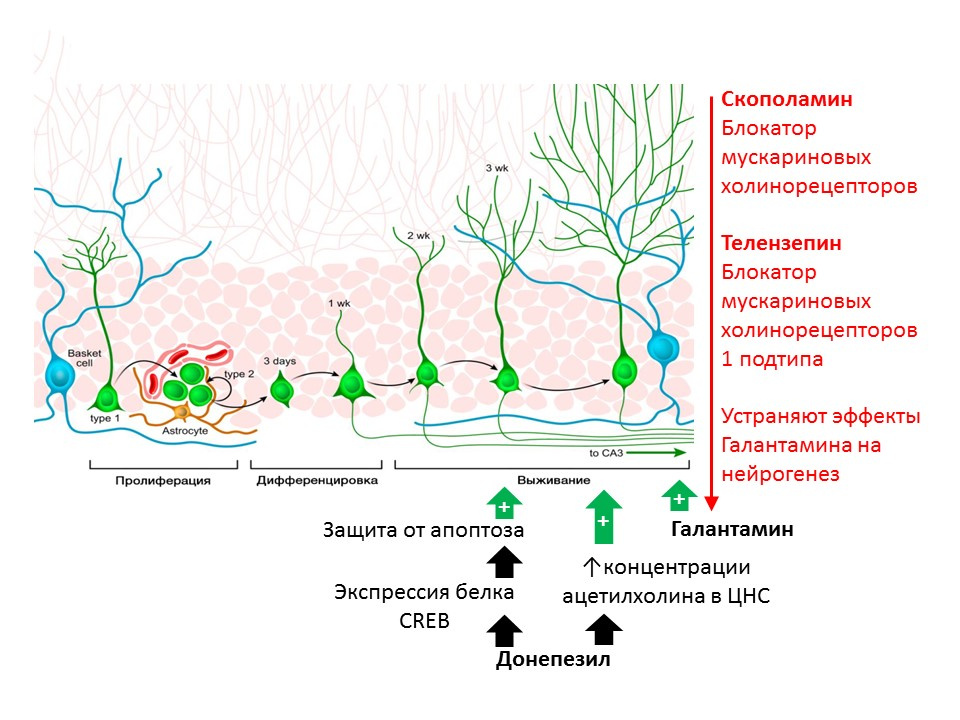 Kotani S等。 H. Donepezil是一种乙酰胆碱酯酶抑制剂,可增强成年海马神经发生。 2008; 175(1-3):227-30
Kotani S等。 H. Donepezil是一种乙酰胆碱酯酶抑制剂,可增强成年海马神经发生。 2008; 175(1-3):227-30
Kita Y等。 加兰他敏通过鼠中的毒蕈碱和α7烟碱样受体促进成年海马神经发生。 Int J Neuropsychopharmacol。2014; 17(12):1957-68。在使用防止其破坏的物质(乙酰胆碱酯酶抑制剂-一种分解ACX的酶)中,乙酰胆碱(ACX)的调节作用在大鼠中显示出来。 多奈哌齐和加兰他敏增加中枢神经系统中ACX的浓度(多奈哌齐还增加CREB蛋白(一种转录因子)的表达,从而增强对未成熟神经元免受凋亡的保护)并增加未成熟神经元的存活率。 加兰他敏对神经发生的积极作用被东pol碱(毒蕈碱胆碱能受体的所有亚型的阻滞剂)和telenzepine(毒蕈碱胆碱能受体的亚型1的阻滞剂)消除了,这表明ACh通过激活1型毒蕈碱受体激活了ACh对神经发生的作用(
Kotani S et al。 Chem Biol Interact。2008; 175(1-3):227-30; Kita Y等人,加兰他敏通过小鼠中的毒蕈碱和α7烟碱样受体促进成年海马神经发生。Int J Neuropsychopharmacol。2014; 17(12):1957-68。 )。
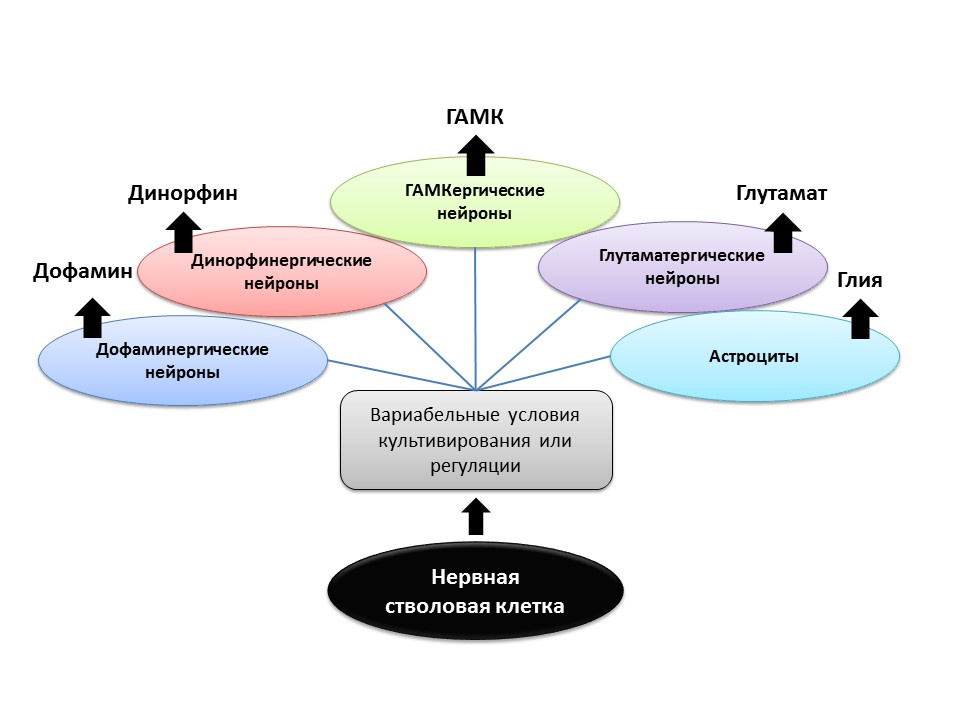
资料来源谢蒂AK。 来自胚胎第19天大鼠海马CA3区的祖细胞在体外产生区域特异性神经元表型。 海马 2004; 14(5):595-614
Gómez-LiraG等。 海马颗粒细胞的编程和诱导表型。 神经科学杂志。 2005; 25(30):6939-46
Nakamichi N,Takarada T,YonedaY。由γ-氨基丁酸和谷氨酸信号传导介导的神经发生。 J.Pharmacol Sci。 2009; 110(2):133-49
Arenas E.在干/前体细胞中工程化多巴胺能表型:Nurr1,胶质细胞衍生信号和Wnts的作用。 Ann NY Acad Sci。 2005; 1049:51-66
在实验条件下(在动物或细胞培养物中)显示,通过改变调节因子的比例并将神经递质添加至神经干细胞本身,可以对新形成的神经元的神经递质表型(即未来细胞的功能)进行``编程''。 因此,例如,与星形胶质细胞一起培养干细胞可让您获得大量的谷氨酸能神经元(产生谷氨酸的神经元)以及多巴胺-多巴胺能神经元。
将来有可能将这种定向作用用于某种表型的神经细胞的选择性形成。 例如,如果我们知道某种特定疾病丧失了什么类型(帕金森氏病,多巴胺能性疾病死亡,阿尔茨海默氏病,胆碱能性疾病)。
但目前,神经发生的这一方面仍未完全探索,谈论神经发生的程序调节还为时过早(很少有研究)。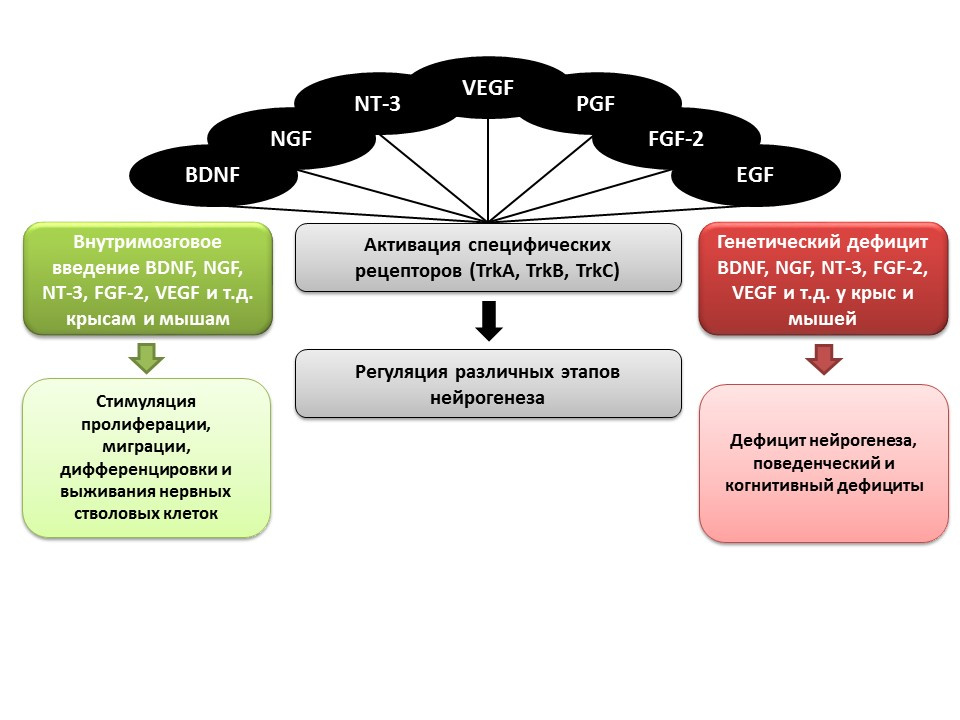
资料来源Pencea V et al. Infusion of BDNF into the lateral ventricle of the adult rat leads to new neurons in the parenchyma of the striatum, septum, thalamus, and hypothalamus. J.Neurosci. 2001;21:6706–6717
Bath KG et al. Variant brain-derived neurotrophic factor (Val66Met) alters adult olfactory bulb neurogenesis and spontaneous olfactory discrimination. J Neurosci. 2008; 28:2383–2393
Gascon E, Vutskits L, Zhang H, et al. Sequential activation of p75 and TrkB is involved in dendritic development of subventricular zone-derived neuronal progenitors in vitro. Eur J Neurosci. 2005;21:69–80
Kahn MA, Kumar S, Liebl D, et al. Mice lacking NT-3, and its receptor TrkC, exhibit profound deficiencies in CNS glial cells. Glia. 1999;26:153–165
Shimazu K, Zhao M, Sakata K, et al. NT-facilitates hippocampal plasticity and learning and memory by regulating neurogenesis. Learn Mem. 2006; 13(3):307-15)
Zhao M, Li D, Shimazu K, et al. Fibroblast growth factor receptor-1 is required for long-term potentiation, memory consolidation, and neurogenesis. Biol Psychiatry. 2007;62:381–390
如前所述,生长和神经营养因子在神经发生调节中起着重要作用,包括:- 脑神经营养因子(BDNF)
- 神经生长因子(NGF)
- Neurotrophin-3(NT-3)
- 血管内皮生长因子(VEGF)
- 血小板生长因子(PGF)
- 成纤维细胞生长因子2(FGF-2)
- 表皮生长因子(EGF)
在动物(大鼠和小鼠)的实验中,显示出上述因子的脑内给药增强了神经发生的所有阶段。先天性缺陷(具有相关因子基因敲除的转基因小鼠)导致神经发生受损,以及行为和认知缺陷。这些数据证实了这些因素在神经发生中的重要调节作用。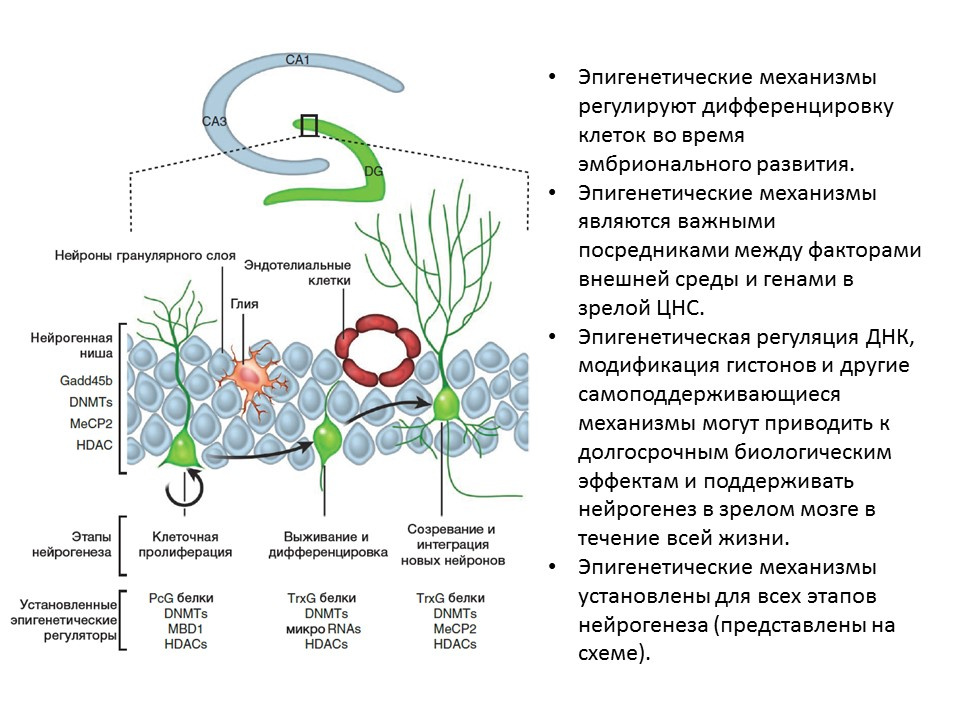 Ma DK,Marchetto MC,Guo JU,Ming GL,Gage FH,Song H.成年哺乳动物脑中神经发生的表观遗传编排。Nat Neurosci.2010; 13(11):1338-44。已经为神经发生的所有阶段建立了表观遗传调控因子,例如:
Ma DK,Marchetto MC,Guo JU,Ming GL,Gage FH,Song H.成年哺乳动物脑中神经发生的表观遗传编排。Nat Neurosci.2010; 13(11):1338-44。已经为神经发生的所有阶段建立了表观遗传调控因子,例如:- 组蛋白脱乙酰基酶(HDAC)
- DNA甲基转移酶(DNMT)
- 微小RNA
- 和其他。
资料来源Coras R, Siebzehnrubl FA, Pauli E, et al. Low proliferation capacities of adult hippocampal stem cells correlate with memory dysfunction in humans. Brain. 2010;133(11):3359-72
Cheyne JE, Grant L, Butler-Munro C, et al. Synaptic integration of newly generated neurons in rat dissociated hippocampal cultures. Mol Cell Neurosci. 2011;47(3):203-14
Hernández-Rabaza V, et al. Neuroscience. 2009;159(1):59-68).
Mochizuki N, Moriyama Y, Takagi N, et al. Intravenous injection of neural progenitor cells improves cerebral ischemia-induced learning dysfunction. Biol Pharm Bull. 2011;34(2):260-5
Sahay A., Hen R. Hippocampal neurogenesis and depression. Novartis Found Symp. 2008;289:152-60
Toni N, Laplagne DA, Zhao C, et al. Neurons born in the adult dentate gyrus form functional synapses with target cells. Nat Neurosci. 2008; 11:901–7
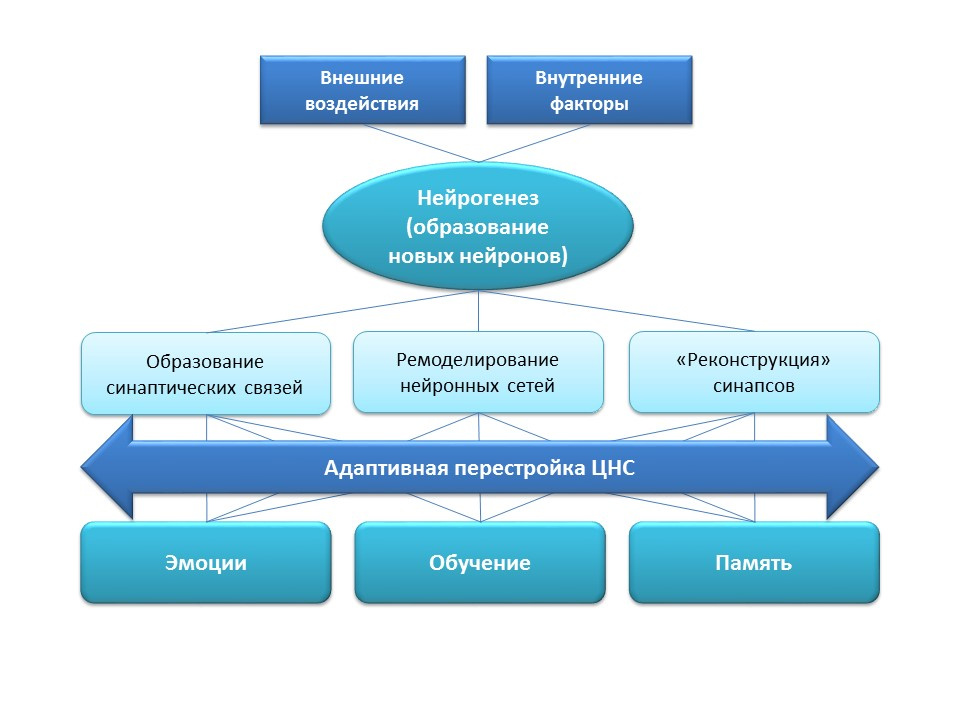
神经发生在中枢神经系统中起着重要的适应性功能,包括新的突触连接的形成(新神经细胞的参与),现有神经网络的重塑(重组),这取决于外部因素(训练,身体活动,压力等)的影响,丢失突触连接的“重建”(在外部和内部因素的影响下)。所有这些适应性变化都直接影响情绪反应,学习过程和记忆。神经发生有助于神经系统在新的条件和任务下保持其“可塑性”,改变和重建。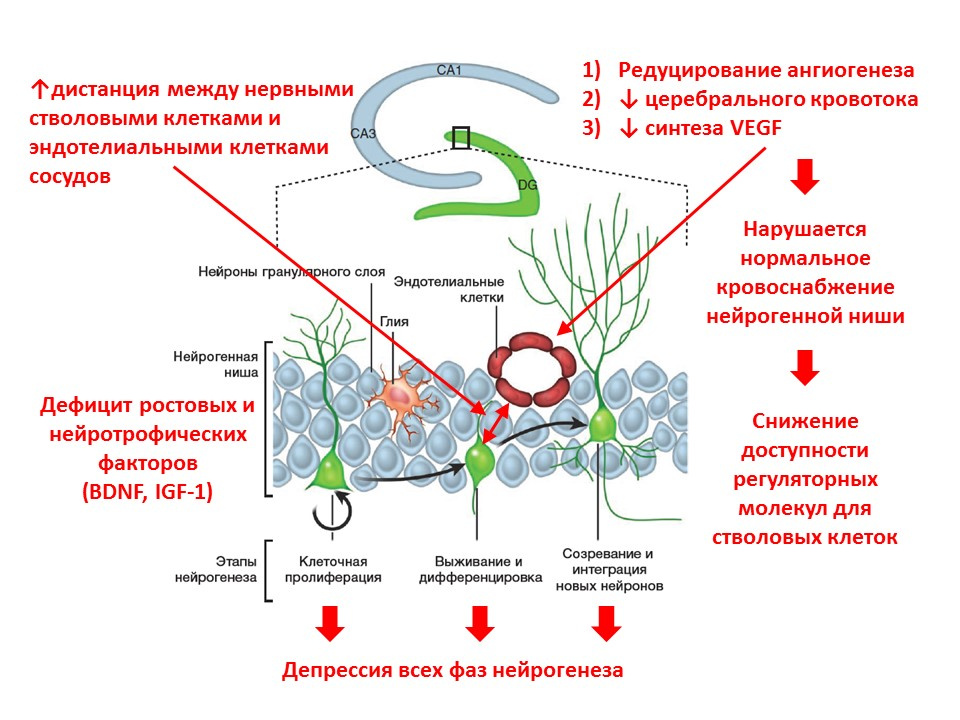
资料来源Lichtenwalner RJ et al. Neuroscienc 2001;107, 603–613
Luo J, et al. Aging Cell. 2006;5(2):139-52
Cuppini R et al. Hippocampus. 2006;16(2):141-8;
Shetty AK, Hattiangady B, Shetty GA. Glia 2005; 51, 173–186
Drapeau E, Nora Abrous D. Aging Cell. 2008;7(4): 569-89
Hattiangady B, Shetty AK. Neurobiol. Aging.2008; 29: 129–177
随着年龄的增长,神经发生的过程开始消失。在分子水平上,衰老伴随着神经发生的以下变化:- 生长因子和神经营养蛋白缺乏-主要调节剂之一;
- 大脑的神经递质平衡发生变化(相反,一些介体的数量减少,另一些介体的数量增加);
- 神经发生各个阶段的强度降低;
- 神经源生态位的血液供应减少(血管生成减少,血管内皮生长因子-VEGF的表达减少),从而减少了干细胞调节因子的可用性;
- 神经干细胞与血管内皮细胞之间的距离增加(正常,干细胞在血管附近转化)。
但是,与年龄相关的神经发生变化不是不可逆的,并且可以刺激适应性脑功能。- , , .. (Jakubs K et al. Inflammation regulates functional integration of neurons born in adult brain. J Neurosci. 2008 Nov 19;28(47):12477-88).
- , . . (Bachstetter AD et al. Fractalkine and CX3CR1 regulate hippocampal neurogenesis in adult and aged rats. Neurobiol Aging. 2011;32(11):2030-44).
该图显示了通过诱导神经炎症和施用分形蛋白来刺激老年大鼠神经发生的信息。 这些数据证明了大脑保留了神经发生所具有的适应功能。
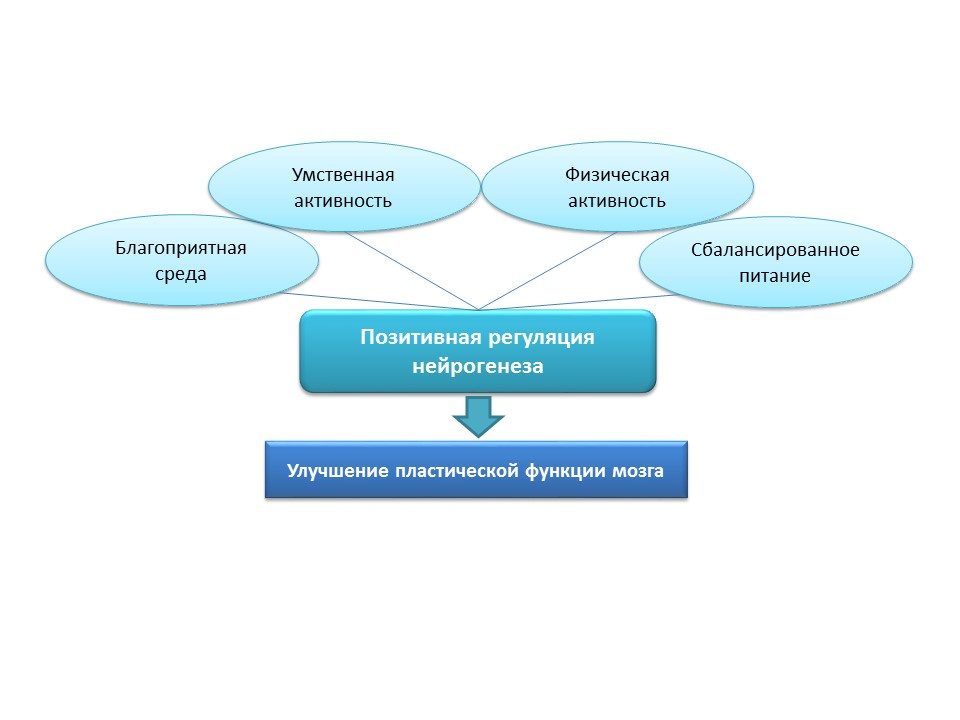
对于老年人自然刺激神经发生,这些策略似乎过于激进,因此,自然积极刺激应被视为“训练”和“支持”神经发生因素:有利的环境,心理活动,体育活动,营养均衡。
 Medvedeva,EV,Dmitrieva,VG,Stavchansky,VV等。 Int J Pept Res Ther(2016)22:197。
Medvedeva,EV,Dmitrieva,VG,Stavchansky,VV等。 Int J Pept Res Ther(2016)22:197。
Jakubs K等。 炎症调节成年大脑中出生的神经元的功能整合。 神经科学杂志。 2008 Nov 19; 28(47):12477-88
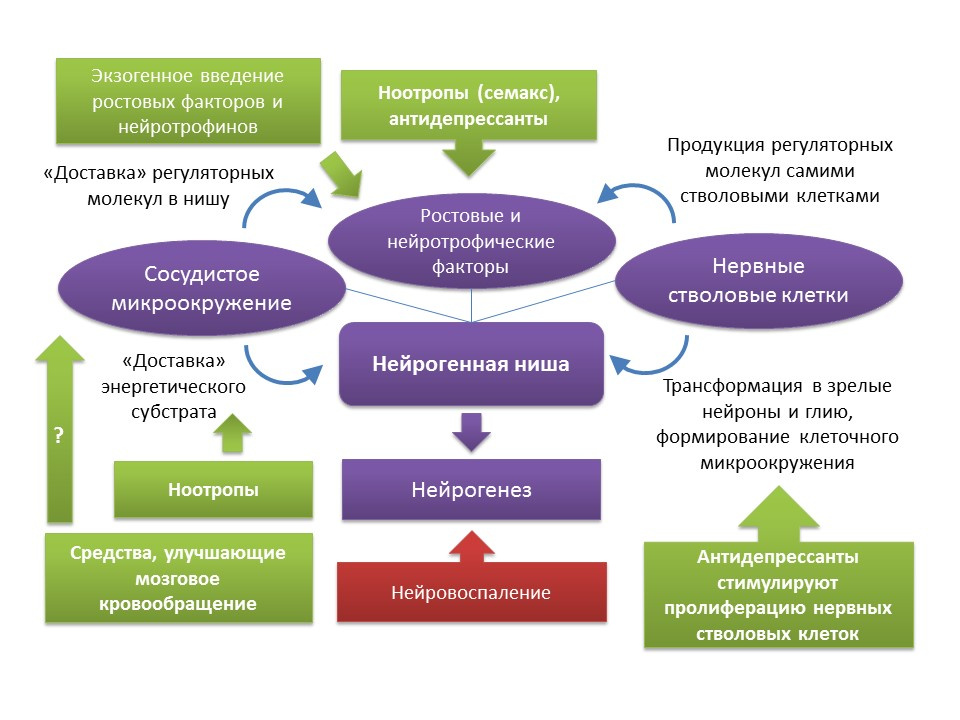
牛磺酸P.促智剂刺激神经发生。 专家观点专家Pat。 2009年5月; 19(5):727-30。如前所述,尽管神经源性利基经历了与年龄相关的变化,但如果有一定的诱因,神经再生功能就可以恢复。 应该注意的是,与此过程相关的衰老速率和与年龄相关的神经发生变化是异质的,并由身体的个体特征决定。 尤其是,人们认为,最初具有较高神经形成水平的人具有较高水平的大脑神经可塑性,因此对中枢神经系统中与年龄相关的变化具有更高的抵抗力。 但是,目前尚无法对整个神经源利基产生全面影响。 如前所述,神经源利基本身就是一种包括许多要素(血管和细胞微环境,一组生长和神经营养因子)的微环境。 一方面,由于对这种“微环境”(内源性调节分子的引入或诱导;药物刺激新形成的神经元的增殖,分化和存活;刺激中枢神经系统的脑循环和能量代谢)的刺激,在神经发生调节方面有相当扎实的药理经验。 另一方面,众所周知的事实是干细胞本身会产生生长和神经营养因子-BDNF,NGF,VEGF等。(
Guzman R.细胞卒中治疗:从细胞置换到营养支持。心血管病专家评论.2009; 7(10 ):1187-1190 )。 这可能会得出这样的结论:为了恢复神经源性利基的功能,可以使用例如干细胞本身或刺激神经干细胞增殖的药物进行治疗。 如已经提到的,各种药理学组的抗抑郁药就是这种方法。 服用抗抑郁药一个疗程后(老年抑郁症后的常见并发症),老年痴呆症风险降低的临床证据可能间接证实了这一结论。 应当指出,有关抗抑郁药对痴呆症风险影响的数据尚有争议。 因此,发现三环类抗抑郁药的使用与痴呆症的患病风险降低有关,相反,MAO抑制剂(单胺氧化酶),杂环类抗抑郁药和SSRIs(选择性5-羟色胺再摄取抑制剂)可能会增加患痴呆症的风险,或对痴呆症的发展没有任何影响(
Kessing LV,Forman JL,Andersen PK。持续的抗抑郁药是否可以预防重度抑郁症患者的痴呆症?Int Clin Psychopharmacol。2011; 26(6):316-22; Lee CW等人。抗抑郁药治疗和痴呆症的风险: J Clin Psychiatry。2016年1月; 77(1):117-22;测验122 )。 然而,这些临床数据在解释时仍需谨慎,因为这些临床评价未考虑许多因素(患者的吸烟史,饮酒,抑郁症的严重程度和形式,伴随的病理学等)。
可以被认为是潜在神经源性药物的另一类有趣的药物是促智药。 促智药物具有许多特定特征:1)仅在记忆功能性病理的背景下起作用; 2)具有动作的累积性; 3)作用机理和化学结构各不相同; 4)行为类似于精神兴奋剂,但没有内在的副作用; 5)迄今为止,关于促智药的临床疗效尚无令人信服的数据。 人们认为,促智药物对中枢神经系统具有复杂的作用:它们改善大脑循环,能量代谢,并与大多数神经递质(GABA,谷氨酸,乙酰胆碱)的受体相互作用。 在实验研究中,已经显示出某些促智剂刺激神经发生的潜在能力。 尤其是,吡乙酰胺增强了人类神经干细胞培养物中的分化过程(
Taupin P.促智药物刺激神经发生。Expert Opin Ther Pat。2009 May; 19(5):727-30 ),另一种促智药物Semax增强了超过20种生长的基因表达。在模拟缺血性中风的条件
下腹膜内给药后大鼠大脑中的神经营养因子和神经营养因子(
Medvedeva,EV,Dmitrieva,VG,Stavchansky,VV等人 Int J Pept Res Ther(2016)22:197 ),这也表明该药物具有潜在的促前体作用。 鉴于对促智药的了解不足,很难谈论它们的神经原活性的具体机制,在这种机制中,它们对中枢神经系统的复杂作用和某些特定成分都可以参与。 显然,这个问题需要进一步的详细研究。
另一类可以刺激神经发生的药物是脑循环调节剂(例如尼莫地平或己酮可可碱)。 然而,关于这些药物对神经发生作用的数据很少。 在大鼠实验中,显示己酮可可碱在脑缺血的情况下改善了短期记忆并降低了海马神经元的凋亡,这可能表明其具有神经保护作用(
Park JH等人, Pentoxifylline减轻了围产期低氧缺血引起的短期记忆障碍抑制大鼠小海马细胞凋亡的作用。Int Neurourol J 2016; 20(2):107-113 )。 相反,尼莫地平显示出抑制小鼠海马中缺血刺激的神经发生的能力(Luo CX等人,L型电压门控钙通道的阻断通过下调成年小鼠的iNOS表达来抑制缺血诱导的神经发生。神经化学杂志,2005年)。显然与它的直接作用机理有关-阻断L型钙通道。
为了获得在与年龄相关的变化条件下神经源性利基的药理操作的完整图片,需要进行更详细的研究,以扩大对相关条件下神经源性药物活性的理解,并使我们得出结论,在开发可预防和纠正与年龄相关的脑神经源性活性下降的药物方面,存在着有希望的方向。
神经发生在中枢神经系统各种病理中的作用
资料来源博德(Bordt),EA(EA),波斯特(Bolster),英国医学(2014)。 NADPH氧化酶和线粒体来源的活性氧物种在促炎性小胶质细胞活化中的作用:两党事务? Free Radic Biol Med,34–46
Liu J,Solway K,Messing RO,Sharp FR。 沙土鼠短暂性整体缺血后齿状回中神经生成的增加。 神经科学杂志。 1998; 18(19):7768-78
Zhang ZG,Zhang L,Jiang Q,et al。 VEGF增强血管生成并促进缺血性脑中的血脑屏障泄漏。 J临床投资。 2000; 106:829-38
中山D,松山T,石桥上田H等。 中风后人大脑皮层损伤诱导的神经干/祖细胞。 Eur J Neurosci。 2010; 31(1):90-8)
Minger SL,Ekonomou A,Carta EM等。 脑梗塞后人脑中的内源性神经发生。 雷根医学。 2007; 2(1):69-74)。
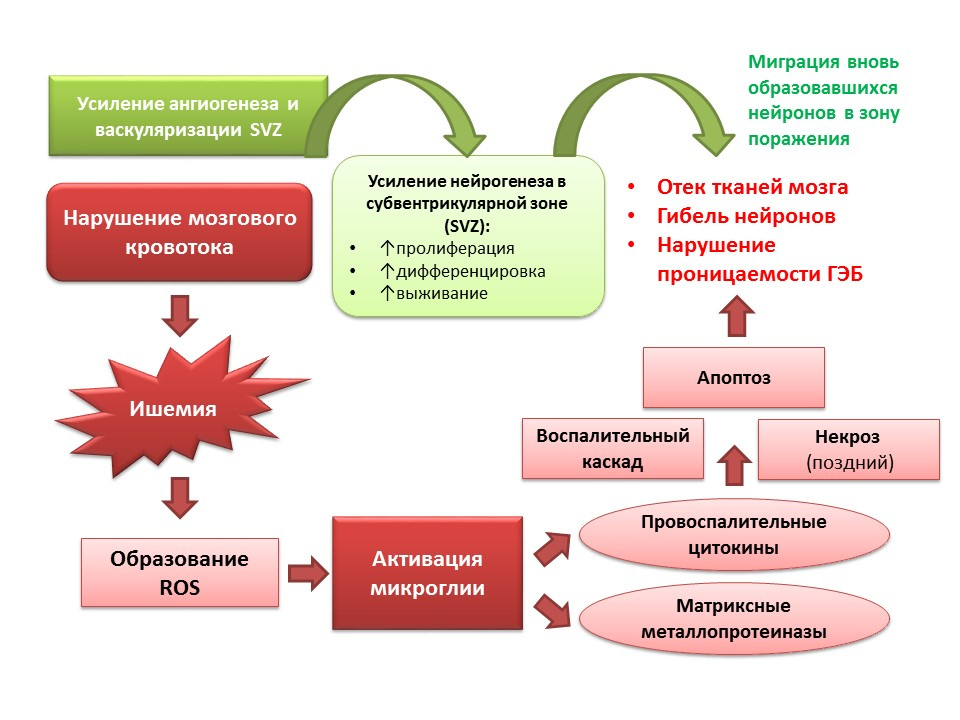
神经发生在CNS病理学的背景下也执行适应性功能。 特别是缺血性中风会导致神经细胞死亡(幻灯片中显示了发病机理的简化方案)。 但是,局部缺血会导致少数神经元快速死亡(数分钟),这就是所谓的局部缺血核。 大多数神经元缺乏葡萄糖和氧气,但它们仍然可以存活数小时(有助于治疗的窗口)-这就是所谓的半影或局部缺血半影。 在缺血性脑损伤的情况下,神经发生在侧脑室的脑室下区被激活,新形成的神经元迁移到半影区。
资料来源Zhang,RL,Zhang,ZG,&Chopp,M.(2013年)。 在缺血性卒中的亚急性恢复治疗中靶向一氧化氮。 关于研究药物的专家意见,22(7),843–851。
陈CM,廖建芳,王玉华,沉玉成。 褪黑激素通过促进缺血性中风小鼠的MT2褪黑激素受体促进内源性神经发生,从而改善神经功能。 免费Radic Biol Med。 2012; 52(9):1634-47
Zhang X,Huang G,Liu H,et al。 叶酸可增强大鼠脑缺血模型中的Notch信号传导,海马神经发生和认知功能。 营养神经科学。 2012; 15(2):55-61
Zhang C,Chopp M,Cui Y,et al。 脑溶素能增强缺血性脑的神经发生,并改善中风后的功能结局。 J Neurosci研究。 2010 Nov 15; 88(15):3275-81
Zhang RL,Zhang Z,Zhang L,et al。 西地那非的延迟治疗可增强老年大鼠局灶性脑缺血后的神经发生并改善其功能恢复。 J Neurosci研究。 2006; 83(7):1213-9
Zheng Z,Chen B.普伐他汀对大鼠脑缺血后神经保护和神经发生的影响。 Neurosci公牛。 2000; 23(4):189-97
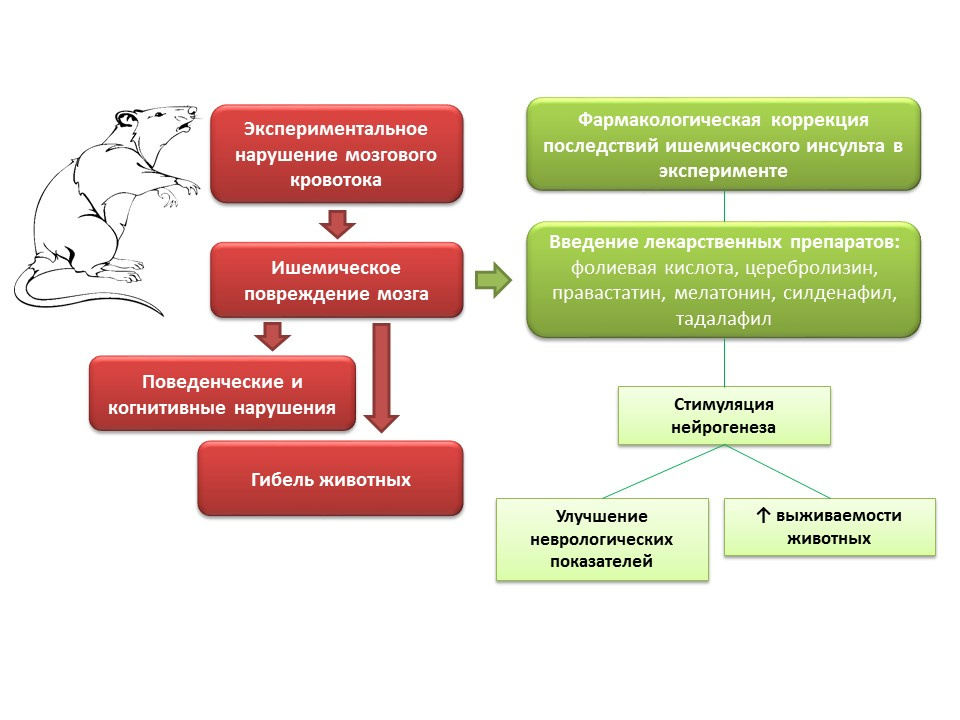
在动物实验中,在对脑血流疾病建模时,会观察到神经系统疾病(认知和行为缺陷)以及大量动物死亡。 向这些动物引入药物具有不同的作用机理,但具有共同的特征-刺激神经发生的能力(叶酸,脑溶素,普伐他汀,褪黑素,西地那非,他达拉非)-改善动物的神经学参数并显着降低死亡率。
操纵缺血性脑损伤神经发生的前景如何?- 有可能使用某些神经保护剂(脑溶素)药物作为急性缺血性脑卒中综合治疗的一部分,以防止半影神经元死亡。
- 对于许多在模拟缺血性中风后已显示出刺激神经发生和改善动物神经状态能力的药物,这种作用不是根本性的,因此,由于其自身作用的存在,其临床使用变得复杂,这可能是不可取的。
- 另一方面,对神经保护作用的潜在机制的研究是一项重要的任务,因为它可以为开发用于缺血性中风的恢复性治疗的根本新型高效药物提供基础。
- 在缺血性脑损伤的背景下,神经发生的药理学调节开辟了创造一组新药的可能性,这些药物可降低缺血性中风后的死亡率并改善患者的生活质量。
资料来源于Y等。 在APP / PS1双转基因小鼠模型中,在阿尔茨海默氏病表型的进展阶段海马神经发生增加。 海马 2009; 19(12):1247-53
Ghosal K,Stathopoulos A,Pimplikar SW。 APP细胞内结构域通过诱导神经炎症而损害转基因小鼠的成年神经发生。 Plos一个。 2010; 5(7):e11866
穆Y,盖奇FH。 成人海马神经发生及其在阿尔茨海默氏病中的作用。 摩尔神经变性剂。 2011; 6:85
Jin K,Peel AL,Mao XO等。 在阿尔茨海默氏病中海马神经发生增加。 Proc Natl Acad Sci美国。 2004; 101(1):343–347
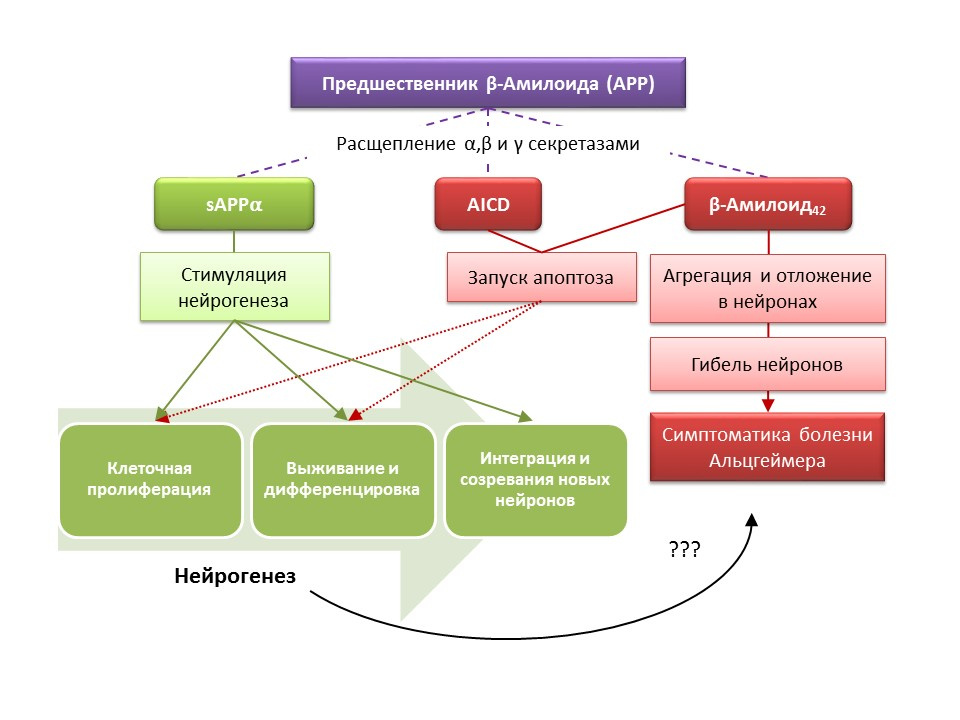
阿尔茨海默氏病(AD)是一种神经退行性疾病,伴有胆碱能神经元(产生乙酰胆碱的神经元)死亡。 目前还不能确切确定阿尔茨海默氏病的病因,该病被认为是多因素的。 AD的发病机理是基于对β-淀粉样蛋白代谢的破坏,β-淀粉样蛋白的各个部分沉积在中枢神经系统神经元中并导致其死亡。 淀粉样蛋白组分对神经发生有不同的影响,因为可溶性淀粉样蛋白-α前体(sAPPα)刺激神经发生的所有阶段,而淀粉样前体(AICD)和β-淀粉样蛋白-42的细胞内结构域诱导神经干细胞凋亡,减少增殖和生存率。 在AD的早期阶段就已经记录了神经发生的激活,那时还没有明显的神经变性和痴呆。 显然,针对AD的神经发生也具有适应性功能。 在大多数AD动物模型(β-淀粉样蛋白代谢受损的转基因小鼠)和来自AD患者的验尸材料中,记录到BrdU阳性细胞数量增加。 但是,有证据表明,在某些AD动物模型和某些AD患者的验尸材料中,神经发生缺乏变化。 最后,关于新形成的细胞替代在AD中死亡的神经元的能力的问题仍然悬而未决。
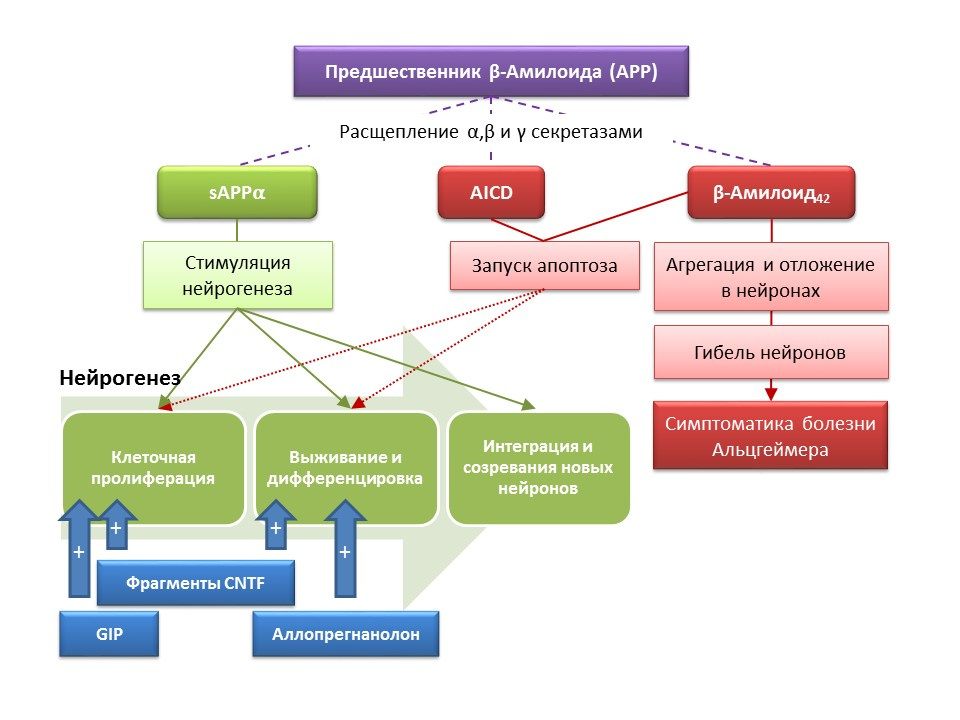 DOI:10.3233 / JAD-2011-110914DOI:10.1016 / j.ejphar.2011.11.007DOI:10.1371 / journal.pone.0024293
DOI:10.3233 / JAD-2011-110914DOI:10.1016 / j.ejphar.2011.11.007DOI:10.1371 / journal.pone.0024293对阿尔茨海默氏病的药理校正新方法进行的实验研究表明,多种物质具有刺激转基因小鼠神经发生的能力(在小鼠中,β-淀粉样蛋白的正常代谢是阿尔茨海默氏病的动物模型),结合了行为行为(定向研究反应,学习和学习)的改善。内存):
- CNTF睫状神经营养蛋白肽6和6A的功能片段的引入刺激了新的干细胞的形成以及新形成的神经元在心室下区,海马齿状回和嗅球中的存活(Rockenstein E,Ubhi K,Doppler E等。神经源性作用的区域比较AβPP转基因小鼠中CNTF衍生肽和脑溶素的测定。J Alzheimers Dis。2011; 27(4):743-52)
- 具有生长因子特性的葡萄糖依赖性促胰岛素肽(GIF)的使用导致海马齿状回中神经干细胞的增殖增加(Faivre E,Hamilton A,HölscherC.比较急性和慢性GIP对认知,突触可塑性和神经发生的影响Eur J Pharmacol。2012; 674(2-3):294-306)。
- 长期和亚慢性给予神经活性类固醇Allopregnanolone有助于新形成的神经元的存活(Chen S,Wang JM,Irwin RW等人.Allopregnanolone在阿尔茨海默氏病的临床前模型中促进再生并减轻β淀粉样蛋白的负担。《公共科学图书馆》,2011年第6期; 8):e24293; Singh C,Liu L,Wang JM,et al.Neurobiol Aging.2012; 33(8):1493-506)
结果表明使用刺激神经发生的物质来减轻阿尔茨海默氏病的症状,并降低神经退行性疾病的进展的前景,这代表了从根本上解决该疾病的药物的新方法。
在阿尔茨海默氏病中控制神经发生的前景- 阿尔茨海默氏病的确切病因至今仍未知,这使得很难找到针对这种病因的新药。
- 在动物阿尔茨海默氏病的遗传建模过程中获得的数据与AD患者的死后物质研究相互矛盾。
- 同时,在阿尔茨海默氏病的动物模型中,已经表明,在疾病的早期阶段(认知功能受损之前)已经观察到神经发生增加。 可以认为这种增强可以被认为是中枢神经系统的适应性保护反应。
- 在阿尔茨海默氏病的动物模型中,已显示出许多物质(睫状中性神经营养蛋白的活性片段-CNTF,葡萄糖依赖性促胰岛素肽-GIP和Allopregnanolone)可以刺激神经发生并改善啮齿动物的行为方式。
- 这些操作的显着缺点是,违反神经发生的事实显然不是阿尔茨海默氏病的病因,而只是症状。 然而,这种疗法的前景可能是减慢疾病的进展并延缓痴呆的发展。
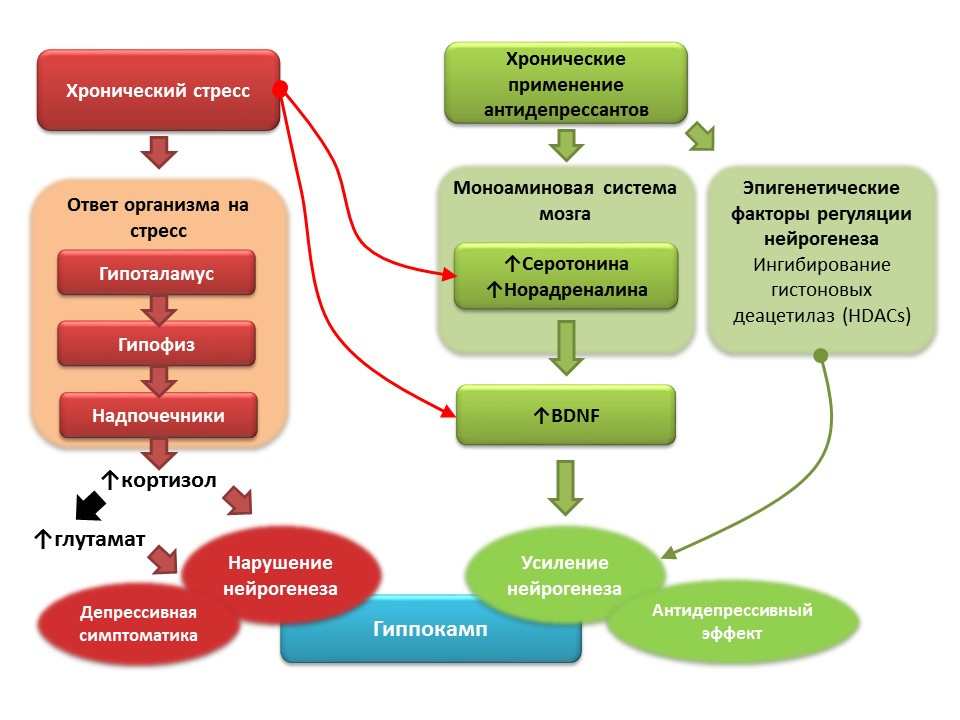 DOI:10.1016 / j.pharma.2013.02.02.006PMID:11124987DOI:10.1186 / s13045-017-0499-7
DOI:10.1016 / j.pharma.2013.02.02.006PMID:11124987DOI:10.1186 / s13045-017-0499-7在临床抑郁症(重度抑郁症)的背景下,还记录了神经生成的减少和海马体积的减少,这些过程在抗抑郁剂疗程后得以恢复。 据信,慢性应激在抑郁症的发病机理中起触发因素的作用,这触发了一系列应激依赖性反应,导致皮质醇的过量产生和中枢神经系统谷氨酸浓度的增加。 这些因素破坏了大脑的可塑性(尤其是神经发生),并导致了抑郁症状的发展。 相反,长期使用抗抑郁药会刺激神经发生,增加中枢神经系统中单胺(5-羟色胺和去甲肾上腺素)(一种脑神经营养因子)的浓度。 一些抗抑郁药还具有调节神经发生的其他机制。 例如,阿米替林能够抑制组蛋白脱乙酰基酶,后者起神经生成的表观遗传调节剂的作用,从而刺激神经干细胞的增殖。
在抑郁症中控制神经发生的前景- 大多数抗抑郁药,无论其作用机理和作用机理如何,都能够在模拟动物模型抑郁症的条件下刺激神经发生。
- 在抑郁症患者中,长期使用抗抑郁药可恢复海马的体积,通常在抑郁症中会减少。
- 所有抗抑郁药的一般作用机理是对单胺系统的影响,特别是增加中枢神经系统中5-羟色胺和去甲肾上腺素浓度的能力。
- 大量实验研究表明,单胺(5-羟色胺和去甲肾上腺素)在调节成年大脑海马神经发生中的重要作用。
- 一个有前途的方向似乎是通过5-羟色胺和去甲肾上腺素调节神经发生的机制的进一步详细研究,对它们的理解可以帮助开发用于抑郁症药理学校正的新药。
- 但是,并非所有抗抑郁药都能单独刺激神经发生,有些只能恢复由慢性压力降低的神经发生的初始水平。
- 因此,另一个重要领域是研究与单胺系统无关的抗抑郁药的机制。例如,抗抑郁药对神经发生的表观遗传调节剂的作用。
- 一个单独的方向是基于脑神经营养因子(BDNF)的制剂的开发以及谷氨酸对中枢神经系统(谷氨酸NMDA受体阻滞剂)作用降低的代表。
研究神经发生的一般观点和方法1.研究调节成熟大脑中神经发生的分子机制。- 成年人大脑中神经发生的调节涉及哪些系统?哪些疾病导致神经发生受到抑制?
- 将这些系统及其各个组件视为神经发生药物校正的潜在目标。
- 寻找可以调节成年大脑神经发生过程的新药物“配方”。
2.进一步的研究应该回答这个问题:在特定的病理过程中,新形成的神经元可以在多大程度上替代丧失的中枢神经系统?3.有必要评估长期外源刺激成人神经发生的可能后果。4.有必要制定一项纠正神经发生疾病的综合策略,其中应考虑所有主要因素:- 血管生成的需要
- 神经发生本身
- 突触发生
- 轴突重塑
- 创造有利的外部条件