
几千年来,人类一直想进入天堂,想要变得像鸟一样。 从达芬奇(Leonardo da Vinci)到莱特兄弟(Wright brothers),许多伟大的思想家设计了各种各样的飞机,其中一些保留在纸上,而另一些则成为现代飞机和直升机的始祖。 现在为一个人飞行并不难-我买了机票,瞧。 我们大多数人甚至都没有想到这种巨大的金属鸟如何将我们从一个大陆转移到另一个大陆。 现代飞机与真正鸟类之间的真相并没有那么多,但是随着自主机器人的发展,情况就略有不同。 许多科学家正在尝试制造机器人鸟类,为了使这项计划取得成功,不仅需要了解鸟类如何飞行,而且还必须了解如何降落。 今天,我们将与您会面进行一项研究,斯坦福大学的毕业生将详细研究种植名叫加里(Gary)及其亲属的鹦鹉的机制。 鸟类如何着陆,他们如何理解它们可以或不能着陆在一个或另一个表面上,以及着陆过程中肢体的运动学是什么? 我们将在研究人员的报告中找到这些和其他问题的答案。 走吧
学习基础
现代飞机和鸟类在着陆方面有什么区别? 例如,波音777需要至少2500 m的跑道,直升机需要平坦(最好)的表面以及直升机。 反过来,鸟类不受种植帆布的几何形状的限制,可以降落在几乎任何表面上(如果我们将鸟类视为一个整体,而不是单个物种)。 如果我们谈论自主机器人,那么它们在着陆方面的能力非常有限。 它们中的一些可以成功地落在特殊设计的表面上,但这不能与普通的啄木鸟相提并论,后者可以在垂直,倾斜,水平的表面上落下,只要有抓爪即可。
 这对我来说很方便(啄木鸟/梅勒甲蚁)。
这对我来说很方便(啄木鸟/梅勒甲蚁)。研究人员指出,他们的前任已经进行了一些研究,量化了种植期间鸟类腿部和翅膀的动态。 但是,仍然不清楚这种动力学如何根据用作着陆点的表面的不同类型而变化。 毕竟,不仅着陆区的几何形状很重要,而且表面纹理也很重要。
有趣的是,以前没有对活禽进行过研究;因此,在实践中,不可能完全评估爪子和爪子的抓地力。 这就是为什么有些问题仍未解决的原因:鸟类如何确切地理解着陆点是合适的; 触摸和抓握时四肢的动态情况如何; 为什么鸟类在各种表面上着陆是一件小事,等等。
在我们今天正在考虑的工作中,研究人员决定通过观察Lesson的虎皮鹦鹉(
Forpus coelestis )来测试树鸟着陆的机制和动力学。
 课的伙伴。
课的伙伴。受试者自愿降落在具有各种几何形状和纹理的电线杆上。 总共有9个选项:3个“自然”,3个直径更改和3个纹理更改。
之所以这样称呼“自然”选项,是因为这些栖息地是由三棵不同树木的真实树枝(直径19毫米)制成的:
- 加州橡木-一根树枝的不同部分表面粗糙;
- 木棉壮丽-表面相对光滑,树枝柔软;
- 桂花-表面纵向粗糙,树枝坚硬。
对于杆直径经过修改的实验,使用不同直径的普通桦木销作为材料:
- 38毫米-这种鹦鹉的直径太大;
- 19毫米-鹦鹉可以部分抓握(前后手指不碰);
- 6毫米-鹦鹉可以完全抓握(前后手指触摸)。
对于具有不同纹理的实验,将直径19毫米的桦木杆包裹在三种不同的材料中:
- 泡沫-柔软而有弹性的质地;
- 铁氟龙-湿滑的质地;
- 砂纸质地粗糙。
鹦鹉正在测试着陆点的所有9个选项。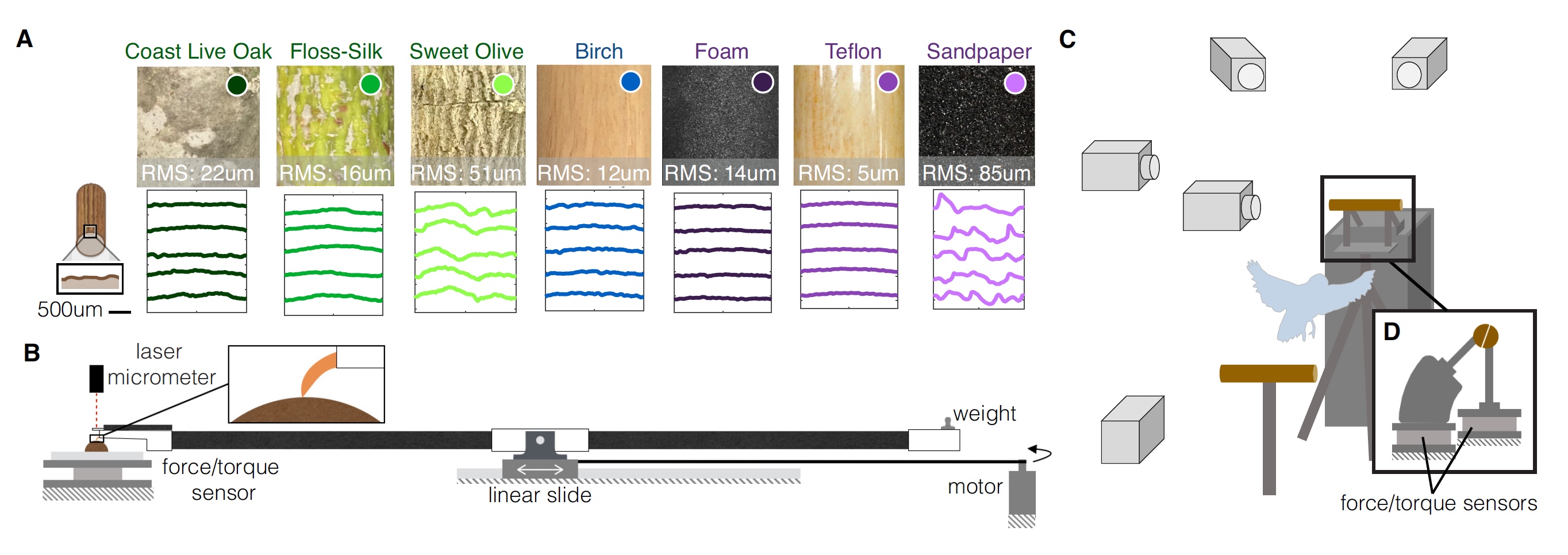 图片编号1
图片编号1对每个穿孔变体进行了三维结构光扫描,这使得可以定量描述每个穿孔变体的表面纹理并重新创建表面轮廓(
1A )。 为了评估这些纹理特征对鹦鹉栖息处和脚部相互作用的影响,研究人员对手指和爪子进行了阻力测试,以测量每个表面的摩擦力。 还评估了爪压以测量表面变形(
1B )。
在图
1C中,我们看到所有测试都是在几台高速摄像机的注视下进行的,并且将杆子锯成两半。 力传感器和力矩分别安装在每个传感器上。
研究成果
首先,考虑降落期间机翼和腿部的动态。
在所有着陆面变体的所有测试中,受试者均表现出相同的经典动态。 着陆始于机翼制动(空气阶段),此后,爪子吸收与鲈鱼接触的剩余冲动(吸收阶段)。 两只爪子之间的接触延迟了几毫秒,即 与杆接触的第一只脚是首选脚:
- 第1个主题-在100%的情况下为右爪;
- 主题2-在83%的情况下为左脚;
- 主题3-在85%的情况下为左脚。
接触后,当鹦鹉将手指更牢固,更牢固地固定在栖息处时,紧固阶段便开始了。 随后是一个阶段,这个阶段并不总是很容易找到,但通常是调整阶段,当阶段以某种方式改变爪子或手指在高位上的位置时。
鹦鹉着陆的所有阶段(对应于2A上的方案)。 图片编号2
图片编号2有时,明显高估或低估了测试条件的鹦鹉没有稍微飞过/飞越着陆点(
2B ),这导致了脚的倾斜角的更大分散,从而建立了静态的抓地力。
很好奇的是,如果支腿(
2B )的强度和方向(
2C )的指标有所不同,则着陆力的平均值对于不同的极点仍然令人惊讶地保持不变。
鹦鹉着陆策略可以解释着陆过程中指示器的差异。 有一种理论认为,鸟类通过视觉评估与地面的接触时间
τ (
t )来控制着陆。 为了成功降落,他们调整了进近速度以保持恒定的
τ (
t )。 如果更详细一点:
τ定义为到高位的距离(
s )除以近似速度(
v )。
如果家禽以恒定减速度(
a )减速,则
τ =
s /
v = 0.5
at 2 /
at = 0.5
t ,在这种情况下
τ (
t )= 0.5。
如果
τ (
t )<0.5,则抑制作用将减小,直到与鲈鱼接触的时刻。 如果0.5 <
τ (
t )<1,则制动将增加,直到鸟类与着陆面发生受控碰撞为止。
给定上述运动学,鹦鹉会保持相对恒定的
τ (
2E )值,该值对应于受控碰撞(
2F )的值。 在具有最小直径(6毫米)的杆的测试中观察到最小平均值
τ = 0.80。
τ = 0.94的最大值是在带有泡沫覆盖极的测试中得出的。 这表明,鹦鹉在接近软着陆表面时会增加制动。
在所有表面类型的所有测试中均存在较高的
τ (即大于0.5),表明鸟类在接触时并未完全着陆。
稍微了解着陆的动态后,我们将继续研究羽毛羽毛朋友的脚和爪的运动学。
考虑到着陆过程中行为的均匀性,无论其表面类型如何,研究人员得出的结论是,鸟类对着陆杆的质地和几何形状的差异的反应完全是用爪子和爪子而不是机翼。 听起来合乎逻辑,不是吗? 但是,令人惊讶的是,这并不完全正确。
脚的运动学显示着陆阶段(
3A ,
3B )的定型观念相似(所有类型的栖息地都相同)。
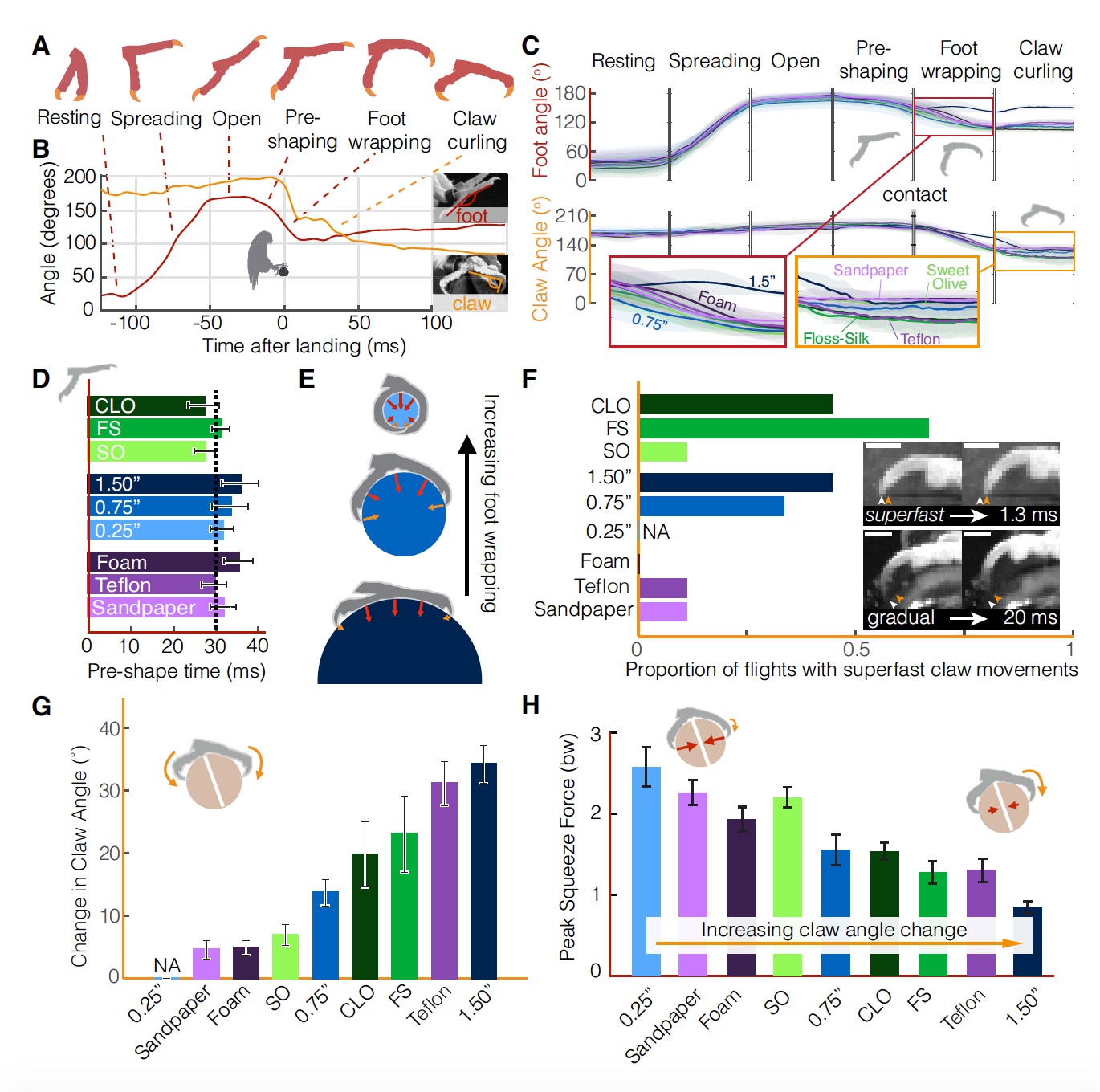 图片编号3
图片编号3飞行过程中,脚处于闭合状态(休息阶段),然后在接触到高位之前大约100毫秒开始张开(持续40±8毫秒的张开阶段),然后在21±7毫秒以内停止时出现“张开”阶段完全公开。
紧接在接触之前,手指开始彼此靠近(捕获的初步形成阶段,持续时间31±10 ms)。 与表面建立接触后,手指开始抓握栖息处(周长阶段,持续时间19±7 ms)。 最后阶段是抓爪,持续185±11 ms。
磁极的纹理和几何形状的差异仅影响着陆表面(
3C )接触后的着陆阶段的运动学。 在所有测试中,即使在预抓握阶段脚开始闭合时,直到接触为止,脚和爪的运动学仍保持不变。
换句话说,如果鸟类对着陆表面的几何形状和纹理的差异做出反应,那么即使在接触之前的各个阶段,即在飞行过程中,也将观察到运动学变化。 仍在空中。 取而代之的是,鹦鹉会在着陆(
3D )之前30毫秒开始预捕获。 这就导致了这样一个假设,即他们对着陆在异常表面上的问题具有可预测的解决方案。
重要的是,在接触之前,保持鸡爪之间的大角度,以防止它们受到伤害。 此外,捕获程度由杆(
3E )的直径确定。 在直径为6 mm的磁极的情况下发生完全捕获,即 鹦鹉可以充分利用其所有手指和爪子的力量来稳定它们。
重要的是要注意,由于光滑的表面,在接触时,爪子和/或爪子可能会滑离两极。 结果,爪角可以变化。 考虑到爪抓取阶段可以持续100毫秒以上,单个爪(
3F )的单个运动可以非常快地(1-2 ms)发生。
研究人员认为,超快速的爪运动可能不仅仅由肌肉收缩来调节。 令人惊讶的是,在脊椎动物中,最快的肌肉运动比这些超快的爪子运动慢5-10倍(例如,人的手臂肌肉-50毫秒,蜂鸟的胸肌-8毫秒)。 由此可以得出结论,鹦鹉爪的运动速度取决于弹性肌腱中存储的能量的释放以及爪的低惯性。 这种快速的爪运动使鹦鹉能够根据着陆表面的几何形状和纹理立即调整抓地力。
至于爪子的捕获程度,取决于杆子的直径和纹理(
3G )。 在直径38毫米的情况下,观察到最明显的抓地力。 值得澄清的是,这仅是捕获爪子,而不是全部腿部。 也就是说,直径越大,就越需要依靠爪子,因为爪子无助(
3C ,
3E )。 相对于表面的爪角在很大程度上取决于其纹理。 例如,在较光滑的表面(特氟龙)的情况下,爪子比在粗糙的表面(砂纸)的情况下“弯曲”更多(
3H ),因为在这种情况下,它们可以简单地稍微压缩表面,从而确保可靠的附着力。 夸张地说,当表面更难压缩时,爪子弯曲得更厉害。
 图片编号4
图片编号4然后研究人员决定从力学的角度检查手指的抗压强度和爪的弯曲度的指示如何变化,以及它们如何影响着陆时鸟类对一个或另一个表面的适应性。
在不同质感的杆上进行的测试表明,指尖(
4A )和爪(
4C )在滑动和抓握时均表现出相似的
剪切力* 。
剪切力* -沿一个特定方向推动身体的一部分,而相反地推动身体的第二部分的不相等的力。
对于爪垫,将剪切力表示为所有表面的法向力的函数的最合适方法是库仑摩擦模型(
4B ),该模型可以预测与干燥表面接触的两个物体之间的摩擦力的方向和大小(因此该模型也称为“干摩擦模型” )
对于爪(
4C ),摩擦系数可以是爪垫(
4A )的摩擦系数的8倍。 除其他因素外,这种差异可能是由爪相对于表面(
4D )的方向引起的:在最柔软的表面上,摩擦力保持相对恒定,但在粗糙的表面上,该力也随着爪角的增加而增加。 但是,即使在相同角度下,测量结果也显示,爪的最大受力系数仍高于护垫(
4E )。
为了补偿离合器附着力的随机性,鸟类可以利用负载平衡和爪在表面上的运动来发挥作用。 多个爪之间的载荷分布有效地缩小了所有爪(
4E )的预期总强度的分布。 此外,鸟类可以张开爪子,这使它们增加了粘附在不平坦表面(如果有)上的可能性。 这增加了摩擦力(
4F )的期望值,这说明了爪子在更难于牢固压缩(
3F )的表面上的弯曲程度更为明显。 在不平整度较小但很大的表面上,拉伸爪子会带来好处。 但是,这种说法与观察结果略有不一致,因为鹦鹉并没有太多地寻求扩大其爪的机会,这表明它们在种植期间没有选择最佳的颠簸。 因此,他们尽可能地弯曲爪,直到达到足够的强度以保持稳定的抓地力为止。 如果在此过程中没有达到此力,则禽鸟将开始调整爪子的位置或起飞。
 图片编号5
图片编号5爪和垫的摩擦系数之间的差异也可以通过这些身体部位的几何形状以及与其接触的着陆表面的几何形状来解释。 摩擦片具有较大的面积和柔软的纹理,可让您将载荷分布在许多接触点,从而产生更均匀的摩擦力。 爪又是粗糙的并且具有较小的面积,因此它们依赖于与表面的几何形状的直接相互作用,即与表面的几何形状直接相互作用。 在准备颠簸。
为了量化几何影响,科学家将矢状面中爪形的宽度定为距爪尖(
5A )的高度的函数。 除了使用现有的表面不规则性之外,爪还可以通过表面变形产生摩擦力。
为了模拟这种相互作用,将爪穿透到各种表面(
5B )中的深度的测量与爪本身的几何形状的测量组合在一起。 通过半径为50μm的球体对爪尖的几何形状进行建模,以承受高达25%的体重(
5C )的载荷。 因此,发现例如,从加利福尼亚橡木到鲈鱼的穿透深度为10μm,即。 爪外圆弧长度的0.2%(约5毫米)。
为了更详细地评估爪子尺寸对牵引力的影响,创建了另一个模型,其中不同尖端直径的爪子附着在不同表面上(
5D )。 不出所料,在没有任何爪子渗入表面的情况下,较粗糙的表面更适合成功捕获(
5E )。
当将表面渗透效果(
5F )添加到模型中时,发现有用的表面增加了。 而且,效果越明显,在垂直轴和爪上的净力矢量之间形成的角度越小。 如果爪尖的半径和质量等距缩放,并且所施加的力取决于质量,则深度系数(穿透深度除以爪尖的半径)也将等距缩放。
由此可以得出一个合理的结论,即该表面上的微小不规则足以使小动物有效地附着在该表面上。
 图片编号6
图片编号6通过将上述所有模型组合在一起,以产生各种作用,力和其他作用,研究人员创建了一个二维模型,该模型同时考虑了爪,爪和整个身体的参数(
6A )。
有关脚与着陆表面的接触力学的数据总量使得可以应用有限的优化来确定鸟类的三维“捕获空间”,以及力和力矩的必要组合(
6B )。 , , : , . , (
6C ).
, . , . , . : , .
, , . , , , . , .
.
结语
, , . . , . , , , . , . - -, - — . , , - , . , , , .
周五顶峰:
鹦鹉不仅飞行完美(如本研究所示,可以降落),而且知道如何玩乐。
谢谢大家的关注,保持好奇心,祝大家周末愉快! :)
, . ? ? 通过下订单或将其推荐给您的朋友来支持我们,
为我们为您发明的入门级服务器的独特模拟,为Habr用户提供
30%的折扣: 关于VPS(KVM)E5-2650 v4(6核)的全部真相10GB DDR4 240GB SSD 1Gbps从$ 20还是如何划分服务器? ( RAID1 RAID10, 24 40GB DDR4).
Dell R730xd 2 ? 2 Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 $199 ! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — $99! . c Dell R730xd 5-2650 v4 9000 ?