
任何系统,无论它多么复杂和多层,都有其自己的基础-没有它的基础将无法以其工作方式运行。 在我们星球的生物圈中,也有基本的砖块,所有东西都可以放在上面。 它们是自养生物-可将无机化合物转化为有机化合物的生物。 今天,我们将与您见面,以色列的科学家在实验室中研究出一种新型的以二氧化碳为食的细菌。 在开发过程中使用了什么方法,细菌如何表现,这项工作对人类意味着什么? 我们从研究小组的报告中了解到这一点。 走吧
学习基础
自养生物可以被称为地球上最古老的生物之一。 据信,第一个自养生物出现在二十亿年前,当时异养(不能从无机物合成有机物)细菌通过进化获得了光合作用的能力。 这个术语本身是在1892年由德国科学家Albert Bernhard Frank提出的。
 阿尔伯特·伯恩哈德·弗兰克
阿尔伯特·伯恩哈德·弗兰克一些生物正处于自养和异养合成的十字路口,因为它们从有机化合物中获得碳,但从无机化合物中获得能量。 按照这种逻辑,自养生物可以分为几个主要的自养生物:光养生物,化学营养生物,放射营养生物,石养生物和混合营养生物。 也有过渡小组,其代表很难归因于合成光谱的一个或另一个边缘,但是它们的分类尚未完成。
顾名思义 ,
光养生物使用光子作为能源或太阳能。 正是这些生物体产生了诸如光合作用的营养。
化学
养分比化学更接近物理。 这类生物利用各种氧化还原反应作为能源,即 化学合成。
研究最
欠缺 ,同时又最奇怪的类型是
放射营养素 -一种称为“蘑菇的放射刺激”现象的结果。 这是由于电离辐射而刺激微观真菌代谢的过程。 1991年,这些生物首次作为黑霉菌的一部分被发现,其样本是从切尔诺贝利核电站获得的。 不久之后(2006年),来自纽约的科学家测试了放射性刺激真菌的假设,发现含有球形黑色素的球形球形小球藻(Cladosporium sphaerospermum)三种真菌,皮氏旺吉氏菌和新隐隐球菌增加了生物量并积累了乙酸盐(乙酸,CH
3 OH) )在辐射水平超出标准500倍的环境中。
岩溶菌通过需氧或厌氧呼吸将无机化合物加工成所需的能量和碳。 只有古细菌(没有细胞核和膜细胞器的单细胞)和细菌的代表才能称得上化学萎缩症。
混合营养者是通用士兵,因为他们可以同时(或根据情况交替使用)几种不同类型的食物,即 例如既是光养生物又是化学营养生物。
我们今天正在考虑的研究的作者认为,对自养生物生命活动过程的更详细的了解将允许大规模应用所学知识。 他们认为,研究自养的最佳方法是创造一种合成的自养生物。 从理论上讲,您可以创建一种以二氧化碳为食的细菌。 但是,将这个过程称为简单语言并没有变成现实。 研究人员自己确定了必须完成的三个主要阶段,才能实现他们的工作。
首先,为了完全过渡到自养营养,身体必须在进入的碳仅由CO
2组成的路径中使用CO
2固定机制,输出分子是进入中心碳代谢并提供所有12种主要生物质前体的有机分子。
其次,人体必须利用酶促机制,通过收集非化学能(光,电等)或通过非碳源的氧化还原过程获得再生能量。
第三,人体必须调节和协调能量收集和固定二氧化碳的方式,以便当二氧化碳是唯一的碳源时,它们共同保持稳定的增长。
以前,曾进行过研究,试图创建一种以CO
2为食的生物,但在这些工作中存在一个大缺陷-体内存在多种碳有机化合物,这些物质可作为营养的“储备”来源。 换句话说,尚不可能创造出仅从CO
2中吸收碳的异养生物。
作为研究的方法论基础,科学家使用
了加尔文循环 (Calvin-Benson-Bassam循环)-植物在光合作用,蓝细菌等中的一系列生化反应。 此循环是最常见的二氧化碳固定机制。
主要的实验是
大肠杆菌 ,我们以“ E. coli”的名字而闻名。
研究成果
首先,必须进行身体的代谢重组和实验室进化,以实现向自养的转变。 考虑了几种候选化合物,它们可以用作固定CO
2的电子供体,从而使细菌转移到完全自养的状态。
选择
甲酸酯*作为电子源,因为该一碳有机化合物可作为还原过程的一部分,但不能自然地支持
大肠杆菌的生长,也不会被生物质吸收。
形成* -甲酸的盐和酯。
甲酸的回收潜力(E
0 = 420 mV)足够低,可以还原细胞中的主要电子载体NAD
+ (
大肠杆菌中 E
0 = 280 mV)。 另一个优点是它可以从可再生资源中以电化学方式获得,而生物质将为负碳。
为了从甲酸盐中收集电子并将其引导至NADH(烟酰胺腺嘌呤二核苷酸)主要细胞能量回收库,即来自甲基营养型细菌
假单胞菌sp的 NAD
+连接的FDH(甲酸酯脱氢酶)。
大肠杆菌中代谢网络的化学计量分析(化学化合物的质量比)表明,在
大肠杆菌代谢网络中添加FDH,Rubisco(核糖双磷酸羧化酶)和Prk(磷酸核糖激酶)足以实现自养生长(下图)。
 图1:实验室修饰的化学营养性大肠杆菌的示意图。
图1:实验室修饰的化学营养性大肠杆菌的示意图。不幸的是,三种重组酶在
大肠杆菌 (BW25113)的主要菌株中的共表达并没有导致自养条件下的生长。 由于化学计量分析未考虑酶的动力学,表达水平和调节水平,因此决定使用自适应实验室进化作为代谢优化工具,以实现自养生长。
该方法是由于以下事实:外源酶机制的异源表达扩大了细胞可能的代谢反应的空间,从而提供了自养生长的可能性。 问题在于不能保证必要的数据流将通过最近扩展的一组反应。
因此,由于
大肠杆菌的中央代谢适合异养生长,因此很可能会使用支持异养生长的流量分配。 这就是为什么要使用实验室进化方法,使其能够沿着所需的代谢途径重定向血流。
实验室进化的最重要过程之一是重排中央代谢以建立对Rubisco羧化流的依赖性,并适应生长培养基以抑制通过原始异养途径的流动(
2A )。 换句话说,有必要利用新陈代谢的异养机制使细菌停止运转,从而转变为自养。
 图2:将异养细菌E. coli转化为化学营养的进化策略的示意图。
图2:将异养细菌E. coli转化为化学营养的进化策略的示意图。首先,在人工进化过程中,排除了在中央碳交换中编码两种酶的三个基因:糖酵解中的磷酸果糖激酶(Pfk)和氧化性戊糖磷酸途径中的葡萄糖-6-磷酸脱氢酶(Zwf)。 第一个具有由两个基因(pfkA和pfkB)编码的两个同工酶。 当细胞在
木糖*上生长时
,这种重排可确保细胞生长取决于Rubisco羧化作用,这是过渡到化学趋化所必需的。
木糖*是戊糖单糖(C 5 H 10 O 5 )。
接下来,进行了Rubisco,Prk,碳酸酐酶(CA)的异源表达,其转化了CO
2 ,碳酸氢根和FDH。 此后是在木糖有限的化学恒温
器*中生长细胞的过程,该化学支持细胞处于恒定碳饥饿状态。
Chemostat *是一种在底物生长的营养培养基中维持最佳平衡和底物浓度时培养微生物的方法。
这种培养基可使细胞增殖(通过细胞分裂使组织生长),但减慢异养
分解代谢途径。
分解代谢* -复杂物质的代谢分解为简单物质或物质氧化(能量代谢)。
细胞生长所在的化学恒温器也含有过量的甲酸盐,并不断用富含CO
2的空气(CO
2含量
为 10%)吹扫。
因此,该生长培养基减慢了异养,导致细胞趋于自养。 实际上,细胞被迫减少对有机糖外部碳贡献的依赖性。
生长方法已经准备就绪,有必要进行验证。 每周一次,从恒化器中取出样品并测试其在自养条件下的生长。 特别地,这些是大肠杆菌的化学-有机营养条件,它由M9型培养基组成,在高CO
2含量(10%)但没有任何其他碳源的气氛中添加30 mM(毫摩尔)甲酸钠。
在化学恒温器中繁殖大约200天(相当于大约150代)后,在缺少木糖的培养基中(即在自养条件下)检测到生长。 该表型存在于当天的所有样本中。 在第350天,将木糖从培养基(
2B )中完全排除。 持续的生长和浑浊意味着在恒化器中仅存在不依赖木糖的细胞。 已经发现,样品需要具有高浓度CO
2的培养基才能生长,这表明碳固定机制。
接下来,科学家选择了生长中最强大的
克隆之一
*进行更深入的分析。 建立
倍增时间* 18±4小时(
2C )。
克隆* -在这种情况下,我们是指一组基因相同的细胞。
时间加倍* -某物尺寸加倍所需的时间。
必须确保生长的细胞确实是自养的,并且在其生长过程中没有“隐藏的”碳源或异养的甲酸活化。 为此,进行了同位素标记的实验。
首先,将进化的克隆在具有
13 C标记的甲酸盐和
13 CO
2的培养基中生长(10代,直到获得稳定的同位素状态)。 接下来,通过液相色谱和串联质谱对
13种 C标记物进行了各种代谢物分析。
13 C * -碳-13,稳定的碳同位素。
代谢物* -任何化合物的代谢产物。
液相色谱和串联质谱法*是一种化学研究方法,将液相色谱分离多种组分的混合物与质谱相结合,可确保各个组分的结构相同。
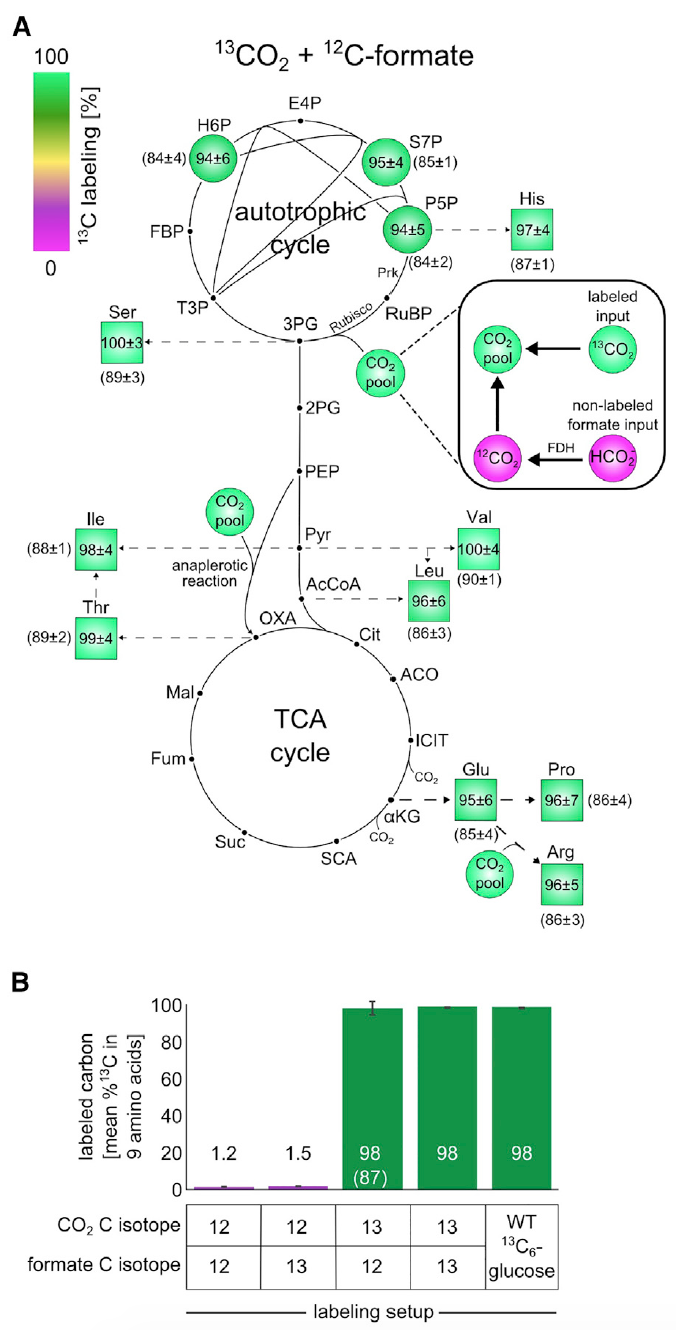 图3:使用13 C的同位素标记实验表明,所有生物质组分均由CO 2作为唯一碳源形成。
图3:使用13 C的同位素标记实验表明,所有生物质组分均由CO 2作为唯一碳源形成。分析表明,中央代谢生物质组成部分中大约98%的碳原子被成功标记。 这些数据对应于标记的甲酸盐和CO
2,其溶解在营养培养基中的含量约为99%的
13 C和1%的未标记的碳酸氢盐。
该观察结果是无可辩驳的证据,表明细胞生物量中的碳正好来自CO
2和甲酸盐。
此外,检查甲酸是否集中在生物质中。 为此,使用碳13标记的甲酸盐使细胞在M9培养基(大气中的CO
2浓度为10%,如先前的实验)中生长。
在给定环境中生长后构建生物质的基础材料的
13 C标记方案显示1-2%(
3B )内的
13 C标记,这是基于天然
13 C含量和少量标记甲酸盐的预期值。 换句话说,结果表明细胞不吸收甲酸盐。
以上实验结果的总体充满信心地表明,生长农作物的碳源仅来自二氧化碳和甲酸盐。 而这又表明经历了实验室进化的
大肠杆菌细胞具有100%的自养能力。
科学家进行了另一项实验来验证这一说法,其中使用了标记为
13 CO
2和未标记的甲酸盐。 由于
13 CO
2的高成本,关闭了进行实验的容器。 这个细微差别非常重要,因为由于封闭的环境(在先前的实验中,容器已通风),由于甲酸盐的氧化而积累的未标记的CO
2得以积累。 这会扭曲观察结果。 但是,可以通过分析标记的谷氨酸盐来监控甚至纠正“污染”过程。
这项经验表明,成功地标记了中央代谢生物质组成部分中约85-90%的碳原子。 从图像
3A和
3B中可以看出,如果我们对
13 C标记的成分进行调整,则生物质中原子的标记将接近100%,这表明
大肠杆菌发展为自养型。
细菌已成为自养生物这一事实是毋庸置疑的。 仍然需要找出在实验室进化过程中发生了哪些遗传变化,即突变。
为了澄清这一点,科学家分离了六个能够在甲酸盐上进行自养生长的克隆,并对它们的基因组和
质粒进行了测序。
质粒*是与染色体物理分离并能够自主复制的DNA分子(基于亲本DNA分子创建两个子DNA分子的过程)。
当木糖仍存在于培养基中(进化第250天)时,分离出两个克隆(克隆1和2),将木糖从化学恒温器的培养基中排除后,分离出三个克隆(克隆3、4和5)(克隆3、4和5)。进化的第400天)。 在一个先前分离的克隆(克隆1)繁殖数个系列稀释周期后,分离出最后一个克隆(克隆6)。
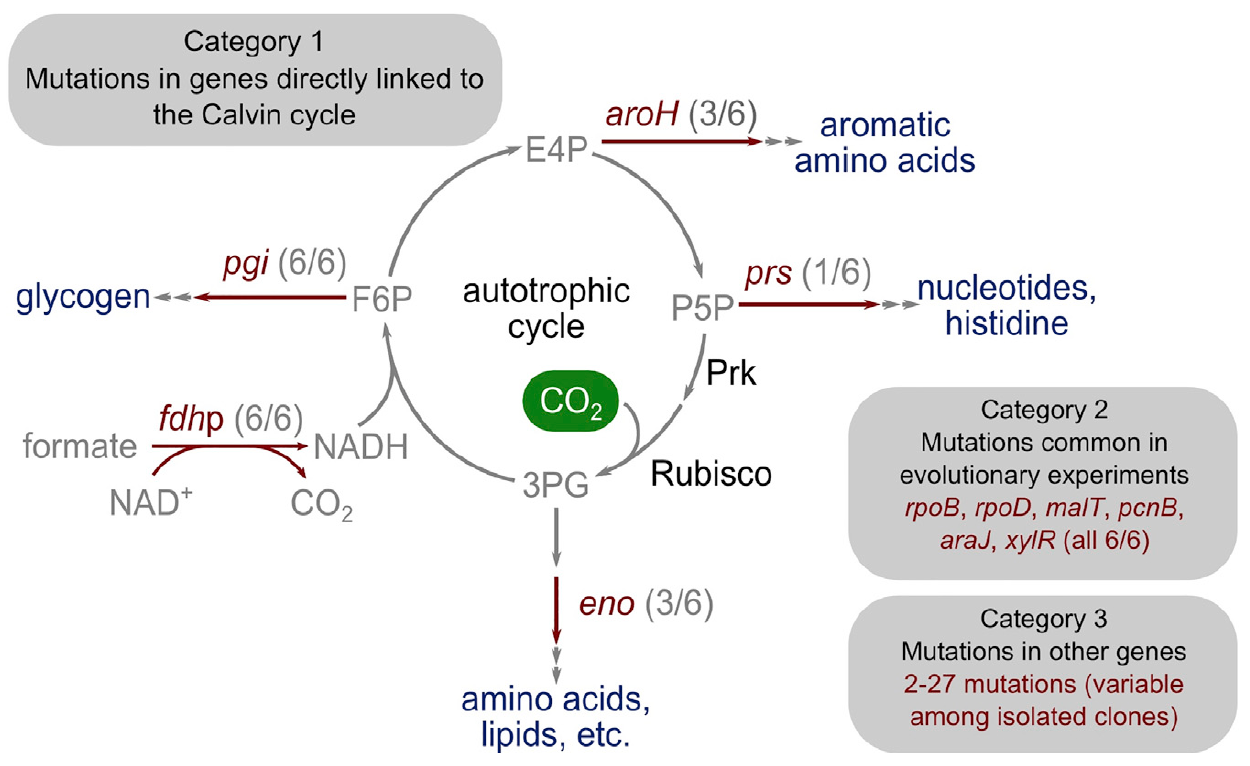 图片4:向自养过渡的遗传基础。
图片4:向自养过渡的遗传基础。令人惊讶的是,突变的数量非常少。 研究人员将它们分为三个主要类别。
第一类由编码具有加尔文循环功能的直接代谢链接的酶的基因组成。 该基因编码核糖磷酸二磷酸激酶,指导核糖磷酸进入生物质。
第二类突变基因包括先前在适应性实验室进化实验中突变的基因:pcnB(R161P),rpoB(D866E),rpoD(F563S),malT(E359K)和araJ(W156)。 科学家将这些突变与实验室进化的整个过程相关联,也就是说,它们不一定与细菌向自养的转变过程相关。 同样,在xylR基因中发现了一个突变,该基因编码负责D-木糖糖(E337K)分解代谢的操纵子的调节蛋白。 它与培养过程中恒化器中木糖的长期饥饿有关,但绝不与自养相关。
第三类突变包括那些没有特征作用的突变,可能是“遗传搭便车”现象的结果。 在不同的分离物中,还有2到27个其他突变基因,其中一些可能是自养表型的突变,但并非严格如此。
将来,科学家打算对基因突变进行其他研究,以确定哪些突变是细菌向自养植物过渡的基础和必要条件。
为了更详尽地了解这项研究的细微差别,我建议您研究一下
科学家的
报告和
其他材料 。
结语
在这项研究中,科学家能够取得出色的结果。 首先,值得注意的是实验室的演变-由科学家控制的过程,使他们能够根据新的“设计”改变人体。
大肠杆菌实际上是通过进化操纵而被迫变成自养生物,吸收CO
2为其自身提供碳。 这样的突变细菌对于遭受全球变暖的社会是极其有用的,其原因之一就是二氧化碳。 , ,
2 , . «» , . -, . -, , ,
2 . , , .
, , , -. , , . , , , .
, , , . :)
一点广告:)
感谢您与我们在一起。 你喜欢我们的文章吗? 想看更多有趣的资料吗? 通过下订单或向您的朋友推荐,
为开发人员提供基于云的VPS, 最低 价格为4.99美元 ,
这是我们为您发明的入门级服务器的
独特类似物: 关于VPS(KVM)E5-2697 v3(6核)的全部真相10GB DDR4 480GB SSD 1Gbps从$ 19还是如何划分服务器? (RAID1和RAID10提供选件,最多24个内核和最大40GB DDR4)。
阿姆斯特丹的Equinix Tier IV数据中心的戴尔R730xd便宜2倍吗? 仅
在荷兰有
2台Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100电视 ! 戴尔R420-2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB-$ 99起! 阅读有关
如何构建基础架构大厦的信息。 使用价格为9000欧元的Dell R730xd E5-2650 v4服务器的上等课程?